

Status epilepticus triggers early and late alterations in brain-derived neurotrophic factor and NMDA glutamate receptor Grin2b DNA methylation levels in the hippocampus
- PMID: 23811393
- PMCID: PMC3830613
- DOI: 10.1016/j.neuroscience.2013.06.029
Status epilepticus triggers early and late alterations in brain-derived neurotrophic factor and NMDA glutamate receptor Grin2b DNA methylation levels in the hippocampus
Abstract
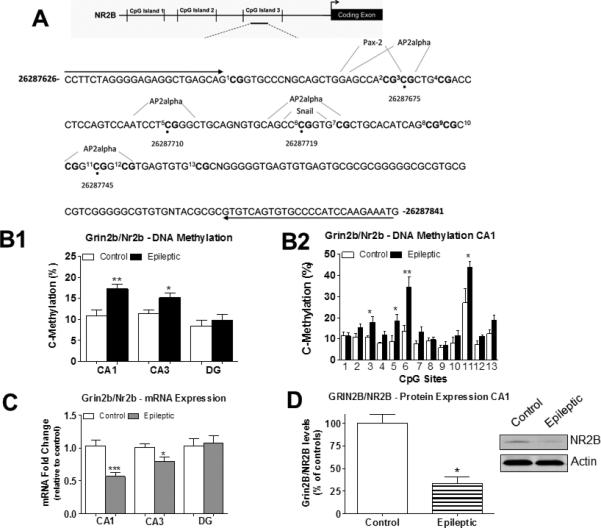
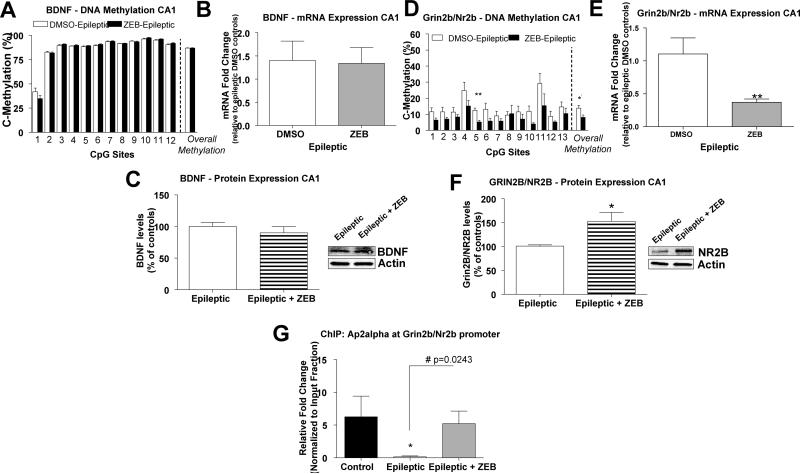
Status epilepticus (SE) triggers abnormal expression of genes in the hippocampus, such as glutamate receptor subunit epsilon-2 (Grin2b/Nr2b) and brain-derived neurotrophic factor (Bdnf), that is thought to occur in temporal lobe epilepsy (TLE). We examined the underlying DNA methylation mechanisms and investigated whether these mechanisms contribute to the expression of these gene targets in the epileptic hippocampus. Experimental TLE was provoked by kainic acid-induced SE. Bisulfite sequencing analysis revealed increased Grin2b/Nr2b and decreased Bdnf DNA methylation levels that corresponded to decreased Grin2b/Nr2b and increased Bdnf mRNA and protein expression in the epileptic hippocampus. Blockade of DNA methyltransferase (DNMT) activity with zebularine decreased global DNA methylation levels and reduced Grin2b/Nr2b, but not Bdnf, DNA methylation levels. Interestingly, we found that DNMT blockade further decreased Grin2b/Nr2b mRNA expression whereas GRIN2B protein expression increased in the epileptic hippocampus, suggesting that a posttranscriptional mechanism may be involved. Using chromatin immunoprecipitation analysis we found that DNMT inhibition restored the decreases in AP2alpha transcription factor levels at the Grin2b/Nr2b promoter in the epileptic hippocampus. DNMT inhibition increased field excitatory postsynaptic potential in hippocampal slices isolated from epileptic rats. Electroencephalography (EEG) monitoring confirmed that DNMT inhibition did not significantly alter the disease course, but promoted the latency to seizure onset or SE. Thus, DNA methylation may be an early event triggered by SE that persists late into the epileptic hippocampus to contribute to gene expression changes in TLE.
Keywords: Bdnf; DNA demethylation; Grin2b/Nr2b; epigenetics; epilepsy; seizures.
Published by Elsevier Ltd.
Figures






References
-
- Amir RE, Van den Veyver IB, Wan M, Tran CQ, Francke U, Zoghbi HY. Rett syndrome is caused by mutations in X-linked MECP2, encoding methyl-CpG-binding protein 2. Nat Genet. 1999;23:185–188. - PubMed
-
- Auzmendi J, Gonzalez N, Girardi E. The NMDAR subunit NR2B expression is modified in hippocampus after repetitive seizures. Neurochem Res. 2009;34:819–826. - PubMed
-
- Bandyopadhyay S, Hablitz JJ. NR2B antagonists restrict spatiotemporal spread of activity in a rat model of cortical dysplasia. Epilepsy Res. 2006;72:127–139. - PubMed
-
- Barreto G, Schafer A, Marhold J, Stach D, Swaminathan SK, Handa V, Doderlein G, Maltry N, Wu W, Lyko F, Niehrs C. Gadd45a promotes epigenetic gene activation by repair-mediated DNA demethylation. Nature. 2007;445:671–675. - PubMed
-
- Binder DK. The role of BDNF in epilepsy and other diseases of the mature nervous system. Advances in experimental medicine and biology. 2004;548:34–56. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
