

Improving photosynthesis
- PMID: 23812345
- PMCID: PMC3729760
- DOI: 10.1104/pp.113.219006
Improving photosynthesis
Abstract
Photosynthesis is the basis of plant growth, and improving photosynthesis can contribute toward greater food security in the coming decades as world population increases. Multiple targets have been identified that could be manipulated to increase crop photosynthesis. The most important target is Rubisco because it catalyses both carboxylation and oxygenation reactions and the majority of responses of photosynthesis to light, CO₂, and temperature are reflected in its kinetic properties. Oxygenase activity can be reduced either by concentrating CO₂ around Rubisco or by modifying the kinetic properties of Rubisco. The C₄ photosynthetic pathway is a CO₂-concentrating mechanism that generally enables C₄ plants to achieve greater efficiency in their use of light, nitrogen, and water than C₃ plants. To capitalize on these advantages, attempts have been made to engineer the C₄ pathway into C₃ rice (Oryza sativa). A simpler approach is to transfer bicarbonate transporters from cyanobacteria into chloroplasts and prevent CO₂ leakage. Recent technological breakthroughs now allow higher plant Rubisco to be engineered and assembled successfully in planta. Novel amino acid sequences can be introduced that have been impossible to reach via normal evolution, potentially enlarging the range of kinetic properties and breaking free from the constraints associated with covariation that have been observed between certain kinetic parameters. Capturing the promise of improved photosynthesis in greater yield potential will require continued efforts to improve carbon allocation within the plant as well as to maintain grain quality and resistance to disease and lodging.
Figures
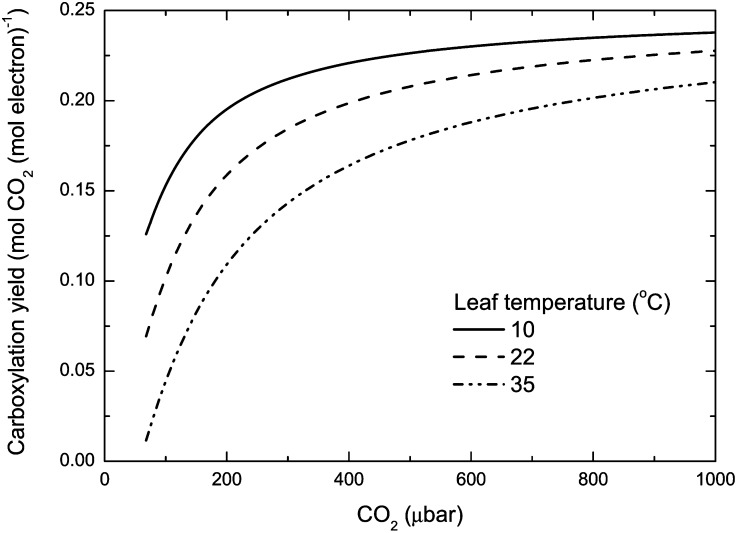
 (Farquhar and von Caemmerer, 1982), where Γ* is the CO2 compensation point in the absence of day respiration (Eqs. 1 and 2). This equation assumes that NADPH regeneration limits photosynthesis. The temperature dependence of Γ* is taken from Brooks and Farquhar (1985).
(Farquhar and von Caemmerer, 1982), where Γ* is the CO2 compensation point in the absence of day respiration (Eqs. 1 and 2). This equation assumes that NADPH regeneration limits photosynthesis. The temperature dependence of Γ* is taken from Brooks and Farquhar (1985).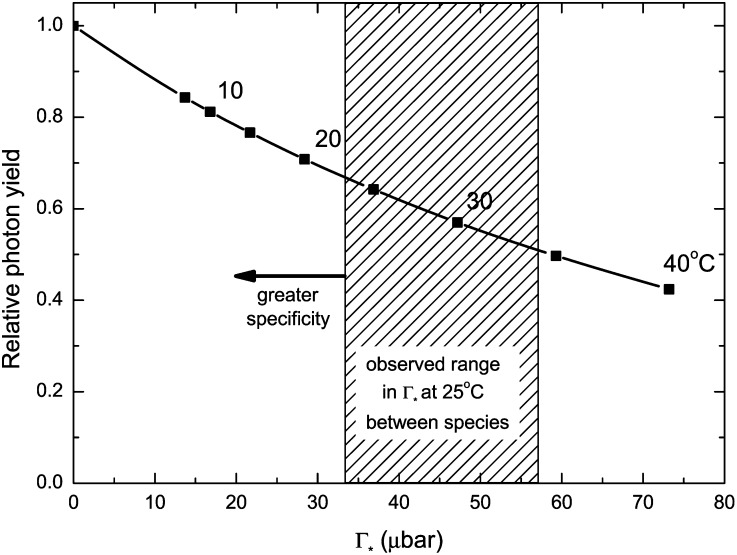
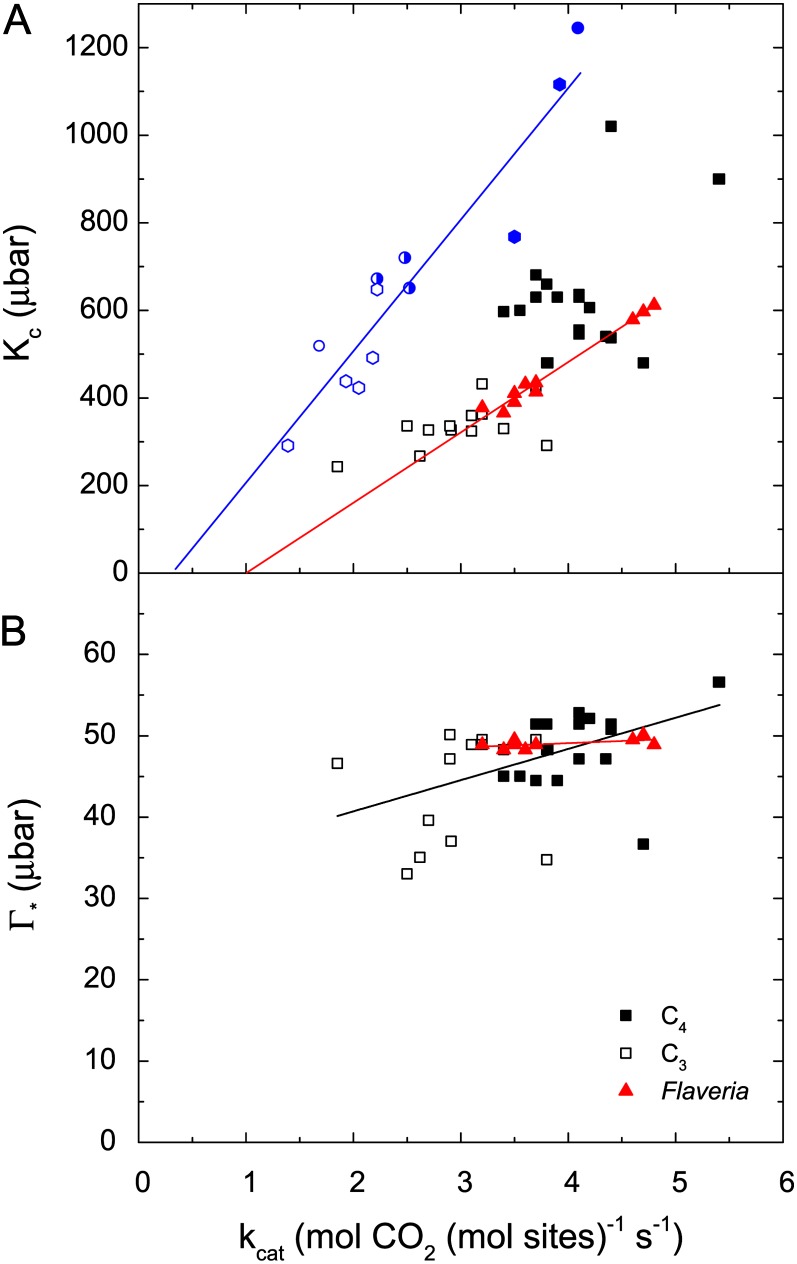
 and a constant value for C are assumed (235 µbar). The temperature response function of Γ* was measured with spinach (Brooks and Farquhar, 1985), and the squares indicate 5°C increments. The striped area illustrates the range in Γ* that has been found for diverse terrestrial plants, including both C3 and C4 species (Kent and Tomany, 1995; Evans and Loreto, 2000; Galmes et al., 2005; Parry et al., 2011). To interconvert between Γ* and Sc/o, divide 3,961 by Γ* or Sc/o (valid for 25°C; von Caemmerer et al., 1994); that is, a value of 40 µbar for Γ* is equivalent to 99 for Sc/o.
and a constant value for C are assumed (235 µbar). The temperature response function of Γ* was measured with spinach (Brooks and Farquhar, 1985), and the squares indicate 5°C increments. The striped area illustrates the range in Γ* that has been found for diverse terrestrial plants, including both C3 and C4 species (Kent and Tomany, 1995; Evans and Loreto, 2000; Galmes et al., 2005; Parry et al., 2011). To interconvert between Γ* and Sc/o, divide 3,961 by Γ* or Sc/o (valid for 25°C; von Caemmerer et al., 1994); that is, a value of 40 µbar for Γ* is equivalent to 99 for Sc/o.
References
-
- Ainsworth EA, Long SP. (2005) What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol 165: 351–371 - PubMed
-
- Amthor JS. (2010) From sunlight to phytomass: on the potential efficiency of converting solar radiation to phyto-energy. New Phytol 188: 939–959 - PubMed
-
- Angus JF, Fischer RA. (1991) Grain and protein responses to nitrogen applied to wheat growing on a red earth. Aust J Agric Res 42: 735–746
-
- Austin R, Bingham J, Blackwell R, Evans L, Ford M, Morgan C, Taylor M. (1980) Genetic improvements in winter wheat yields since 1900 and associated physiological changes. J Agric Sci 94: 675–689
-
- Austin RB, Ford MA, Morgan CL. (1989) Genetic improvement in the yield of winter wheat: a further evaluation. J Agric Sci 112: 295–301
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
