

Control of ribosomal subunit rotation by elongation factor G
- PMID: 23812721
- PMCID: PMC4274944
- DOI: 10.1126/science.1235970
Control of ribosomal subunit rotation by elongation factor G
Abstract
Protein synthesis by the ribosome requires the translocation of transfer RNAs and messenger RNA by one codon after each peptide bond is formed, a reaction that requires ribosomal subunit rotation and is catalyzed by the guanosine triphosphatase (GTPase) elongation factor G (EF-G). We determined 3 angstrom resolution x-ray crystal structures of EF-G complexed with a nonhydrolyzable guanosine 5'-triphosphate (GTP) analog and bound to the Escherichia coli ribosome in different states of ribosomal subunit rotation. The structures reveal that EF-G binding to the ribosome stabilizes switch regions in the GTPase active site, resulting in a compact EF-G conformation that favors an intermediate state of ribosomal subunit rotation. These structures suggest that EF-G controls the translocation reaction by cycles of conformational rigidity and relaxation before and after GTP hydrolysis.
Figures
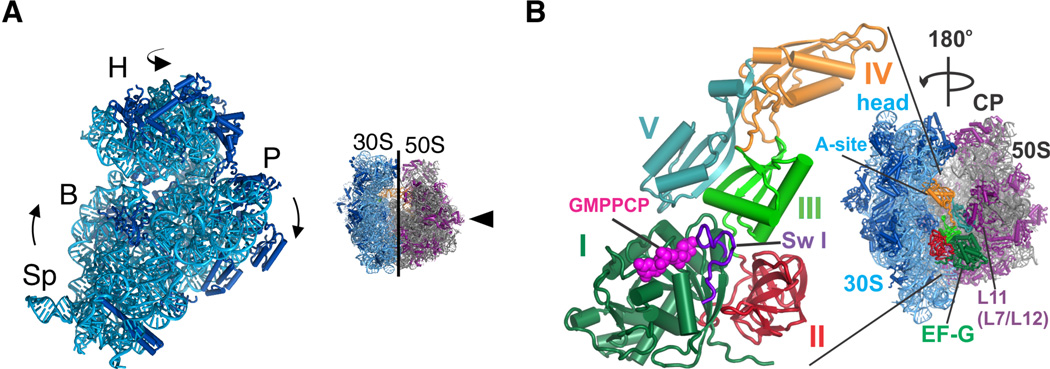
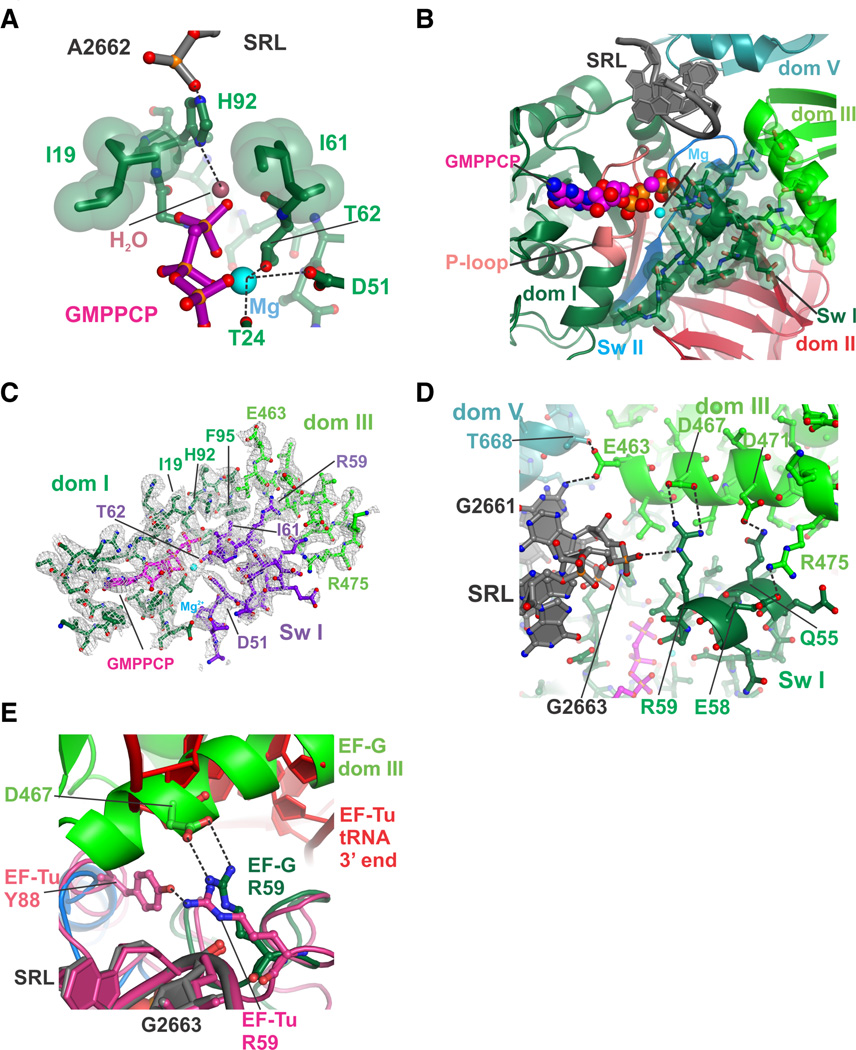
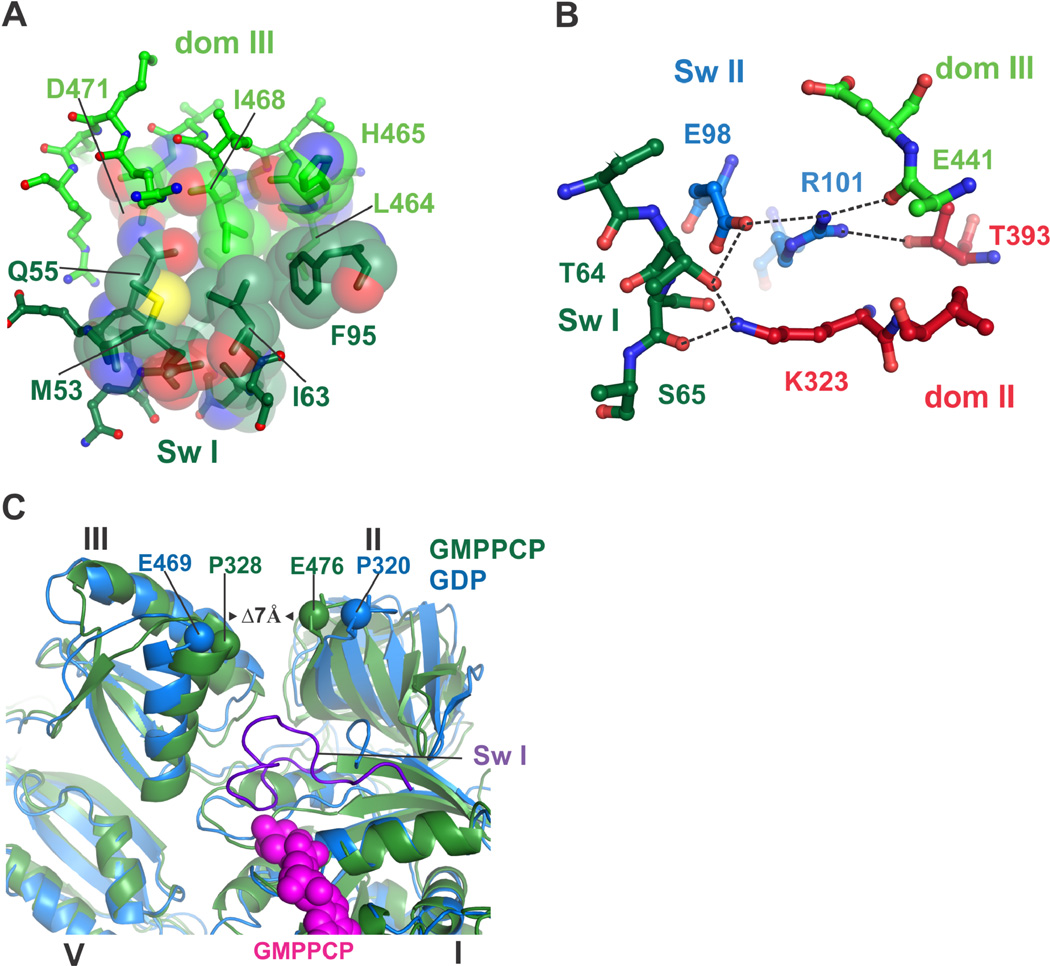
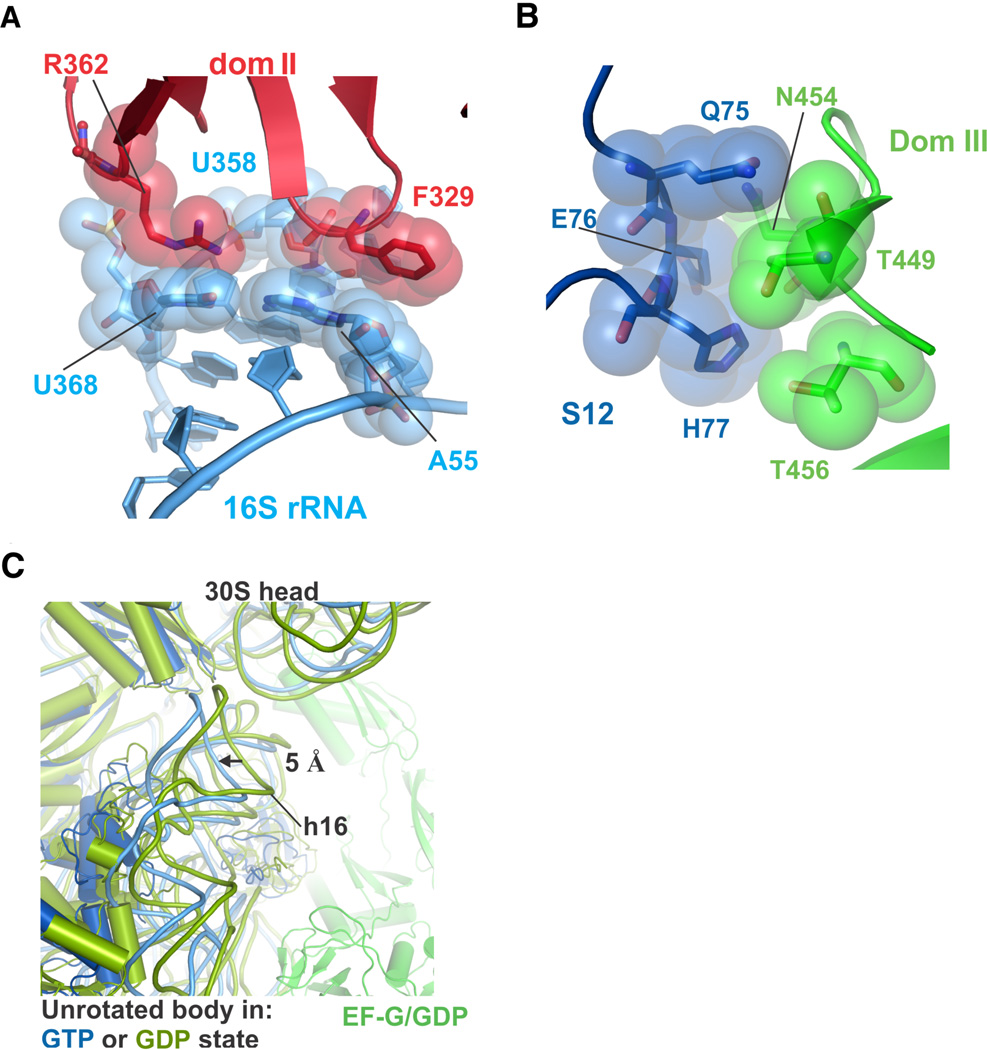
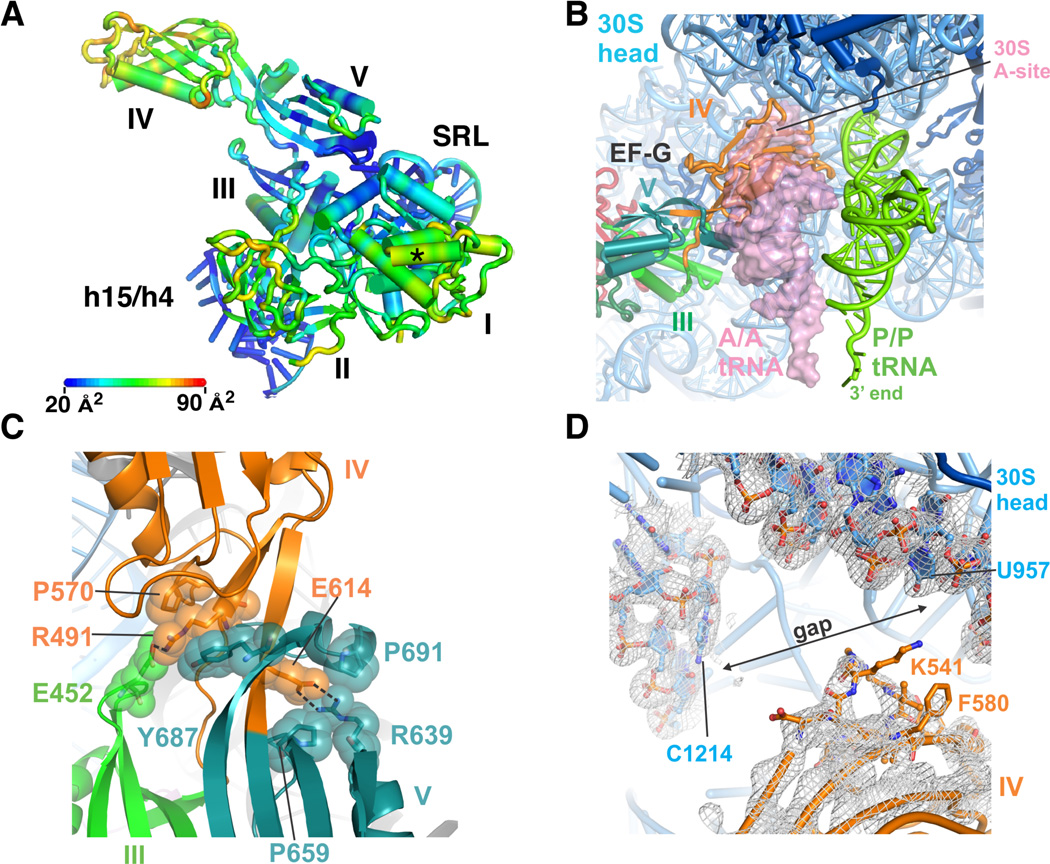
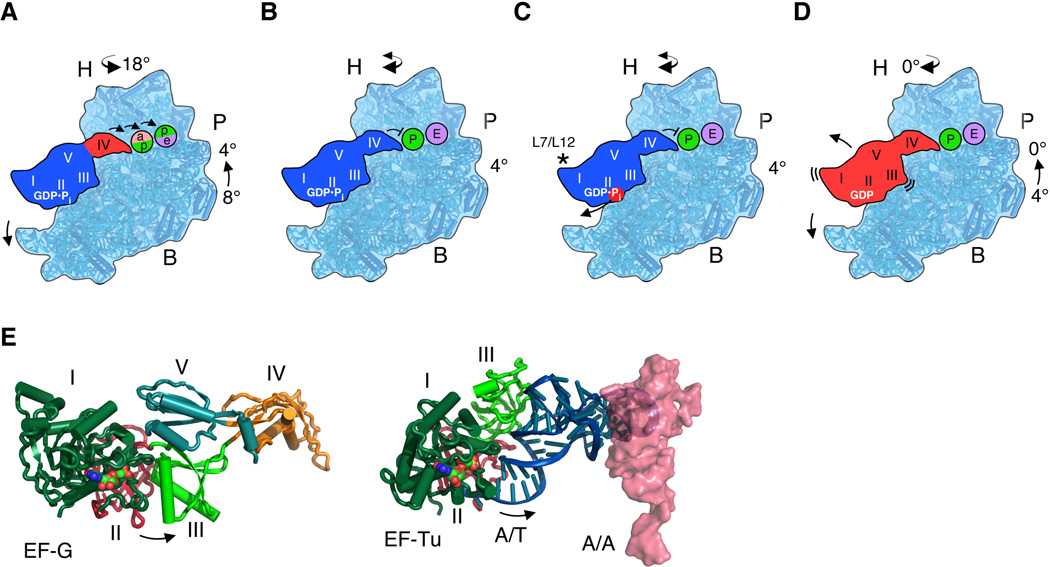
Comment in
-
Biochemistry. Translocation in action.Science. 2013 Jun 28;340(6140):1534-5. doi: 10.1126/science.1240090. Science. 2013. PMID: 23812707 No abstract available.
References
-
- Bourne HR, Sanders DA, McCormick F. The GTPase superfamily: conserved structure and molecular mechanism. Nature. 1991 Jan 10;349:117. - PubMed
-
- Clementi N, Polacek N. Ribosome-associated GTPases: the role of RNA for GTPase activation. RNA Biol. 2010 Sep-Oct;7:521. - PubMed
-
- Sohmen D, Harms JM, Schlunzen F, Wilson DN. Enhanced SnapShot: Antibiotic inhibition of protein synthesis II. Cell. 2009 Oct 2;139:212. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
