

NLRP3 inflammasome activation results in hepatocyte pyroptosis, liver inflammation, and fibrosis in mice
- PMID: 23813842
- PMCID: PMC4008151
- DOI: 10.1002/hep.26592
NLRP3 inflammasome activation results in hepatocyte pyroptosis, liver inflammation, and fibrosis in mice
Abstract
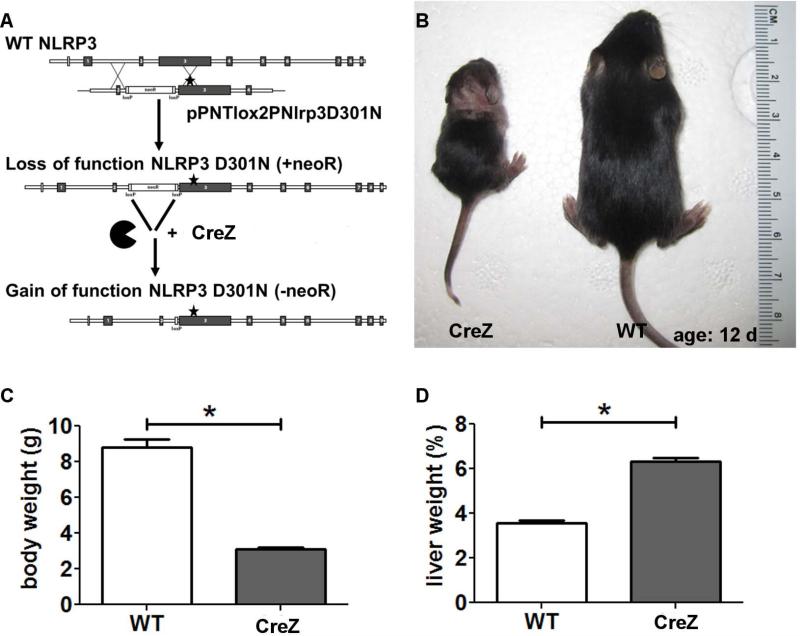
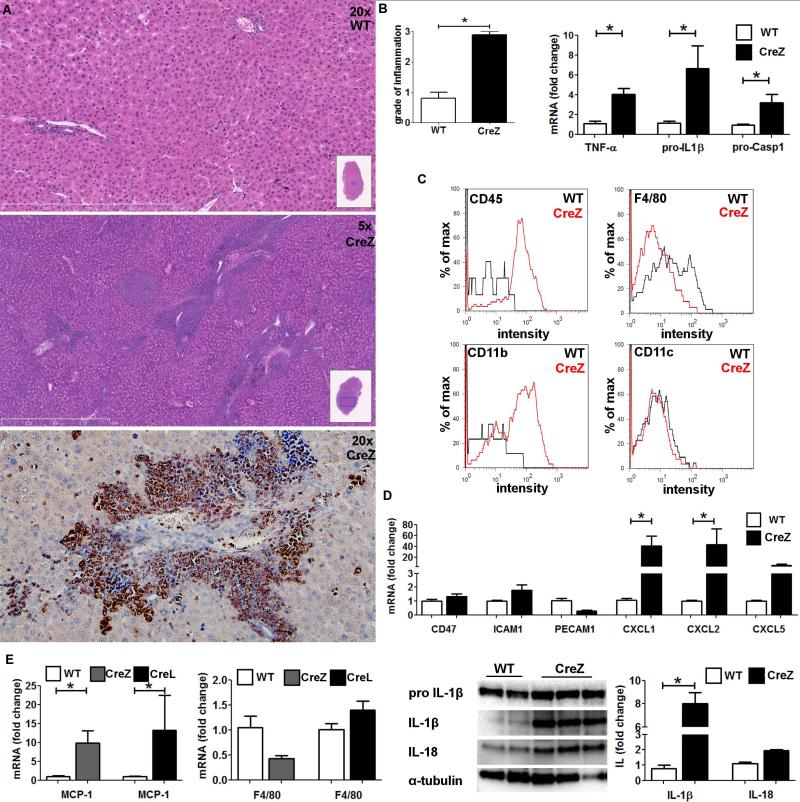
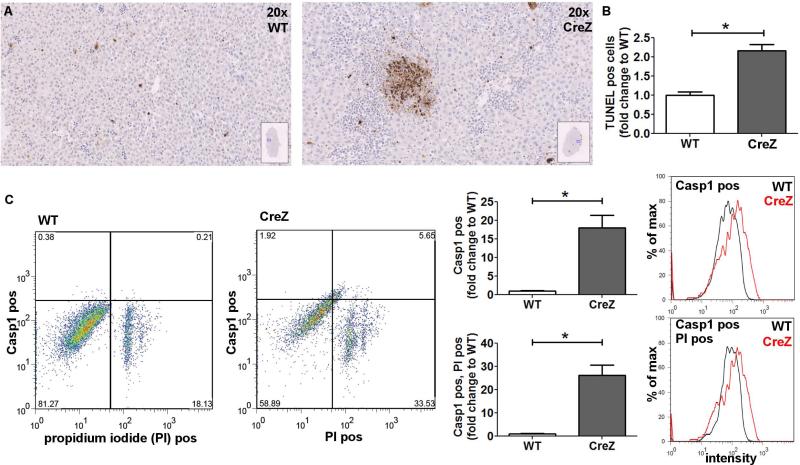
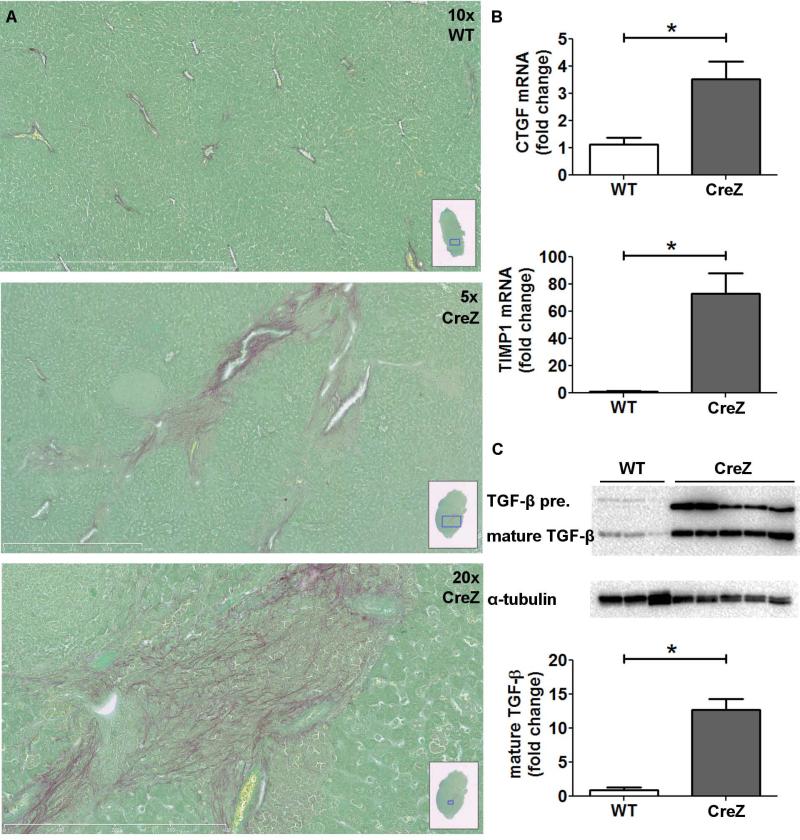
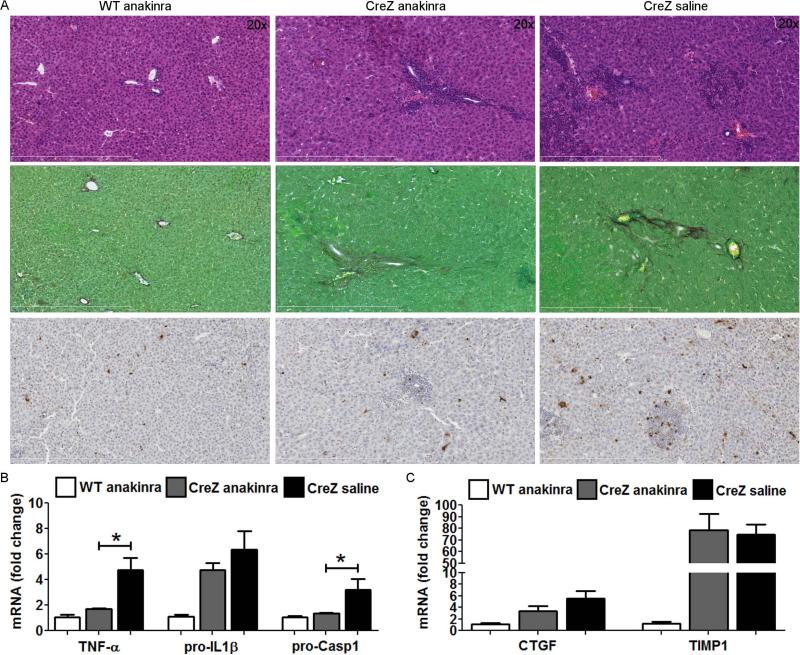
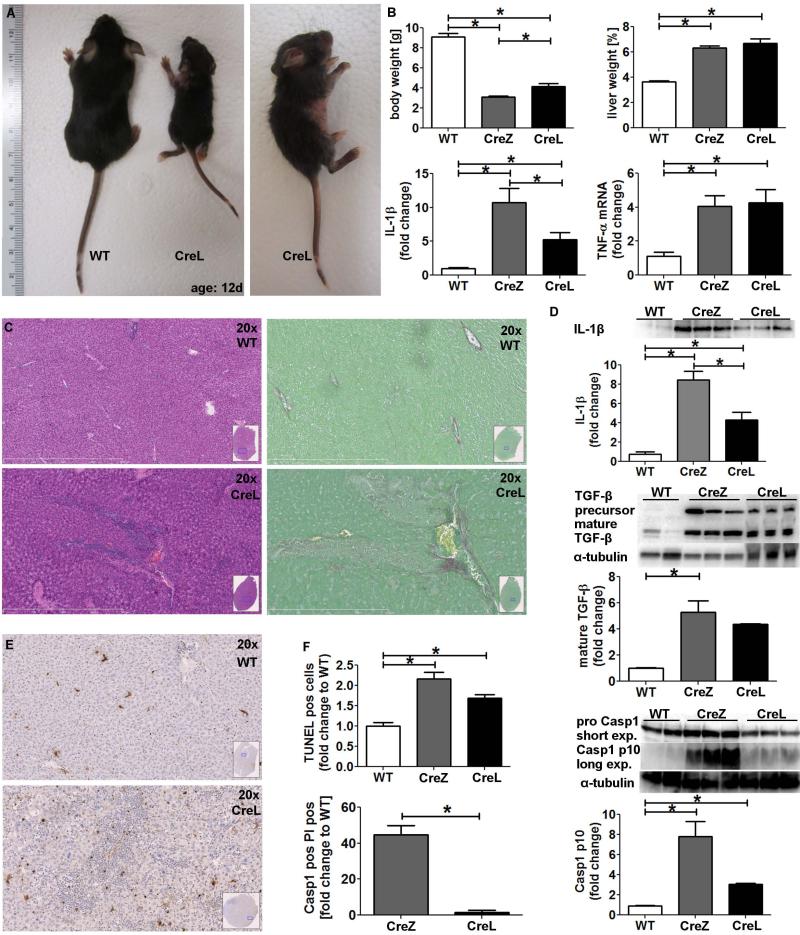
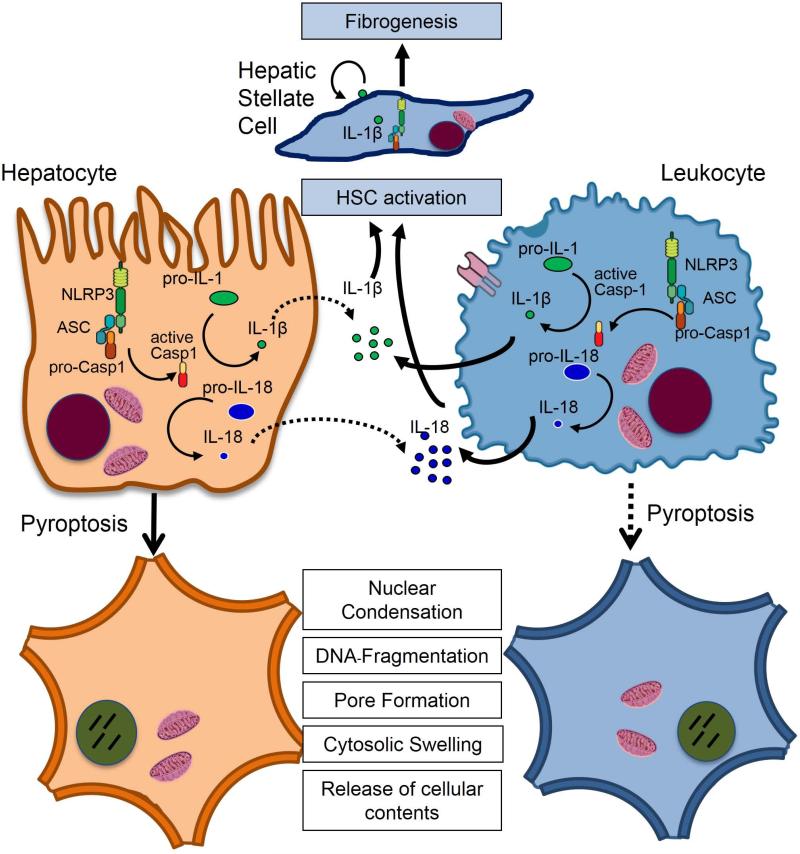
Inflammasome activation plays a central role in the development of drug-induced and obesity-associated liver disease. However, the sources and mechanisms of inflammasome-mediated liver damage remain poorly understood. Our aim was to investigate the effect of NLRP3 inflammasome activation on the liver using novel mouse models. We generated global and myeloid cell-specific conditional mutant Nlrp3 knock-in mice expressing the D301N Nlrp3 mutation (ortholog of D303N in human NLRP3), resulting in a hyperactive NLRP3. To study the presence and significance of NLRP3-initiated pyroptotic cell death, we separated hepatocytes from nonparenchymal cells and developed a novel flow-cytometry-based (fluorescence-activated cell sorting; FACS) strategy to detect and quantify pyroptosis in vivo based on detection of active caspase 1 (Casp1)- and propidium iodide (PI)-positive cells. Liver inflammation was quantified histologically by FACS and gene expression analysis. Liver fibrosis was assessed by Sirius Red staining and quantitative polymerase chain reaction for markers of hepatic stellate cell (HSC) activation. NLRP3 activation resulted in shortened survival, poor growth, and severe liver inflammation; characterized by neutrophilic infiltration and HSC activation with collagen deposition in the liver. These changes were partially attenuated by treatment with anakinra, an interleukin-1 receptor antagonist. Notably, hepatocytes from global Nlrp3-mutant mice showed marked hepatocyte pyroptotic cell death, with more than a 5-fold increase in active Casp1/PI double-positive cells. Myeloid cell-restricted mutant NLRP3 activation resulted in a less-severe liver phenotype in the absence of detectable pyroptotic hepatocyte cell death.
Conclusions: Our data demonstrate that global and, to a lesser extent, myeloid-specific NLRP3 inflammasome activation results in severe liver inflammation and fibrosis while identifying hepatocyte pyroptotic cell death as a novel mechanism of NLRP3-mediated liver damage.
© 2014 by the American Association for the Study of Liver Diseases.
Figures
Comment in
-
Constitutive NLRP3 activation: too much of a bad thing.Hepatology. 2014 Mar;59(3):761-3. doi: 10.1002/hep.26674. Epub 2014 Jan 13. Hepatology. 2014. PMID: 23929761 No abstract available.
References
-
- Martinon F, Burns K, Tschopp J. The inflammasome: a molecular platform triggering activation of inflammatory caspases and processing of proIL-beta. Molecular cell. 2002;10:417–426. - PubMed
-
- Strowig T, Henao-Mejia J, Elinav E, Flavell R. Inflammasomes in health and disease. Nature. 2012;481:278–286. - PubMed
-
- Szabo G, Csak T. Inflammasomes in liver diseases. Journal of hepatology. 2012;57:642–654. - PubMed
-
- Gross O, Thomas CJ, Guarda G, Tschopp J. The inflammasome: an integrated view. Immunological reviews. 2011;243:136–151. - PubMed
-
- Walker NP, Talanian RV, Brady KD, Dang LC, Bump NJ, Ferenz CR, Franklin S, et al. Crystal structure of the cysteine protease interleukin-1 beta-converting enzyme: a (p20/p10)2 homodimer. Cell. 1994;78:343–352. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous