

ARF regulates the stability of p16 protein via REGγ-dependent proteasome degradation
- PMID: 23817020
- PMCID: PMC3748223
- DOI: 10.1158/1541-7786.MCR-13-0207
ARF regulates the stability of p16 protein via REGγ-dependent proteasome degradation
Abstract
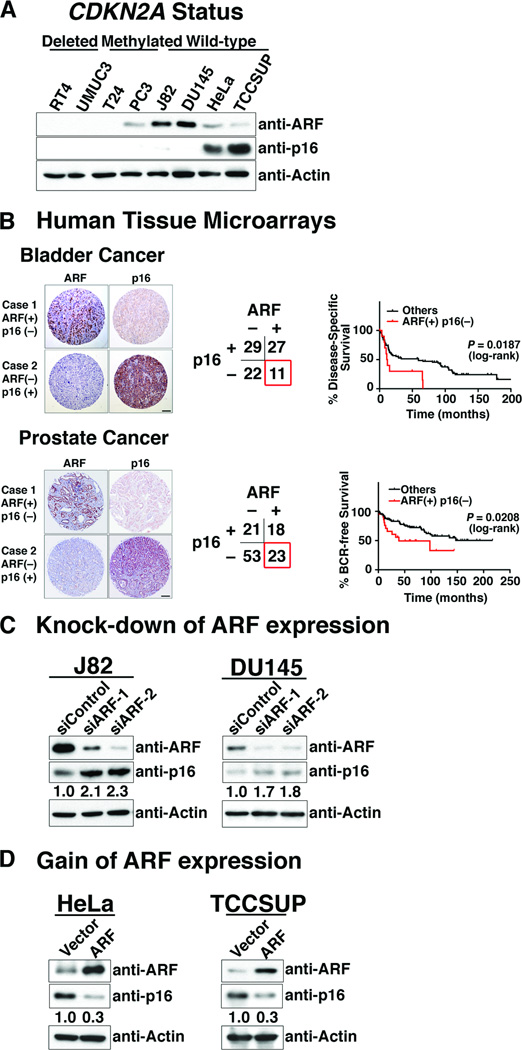
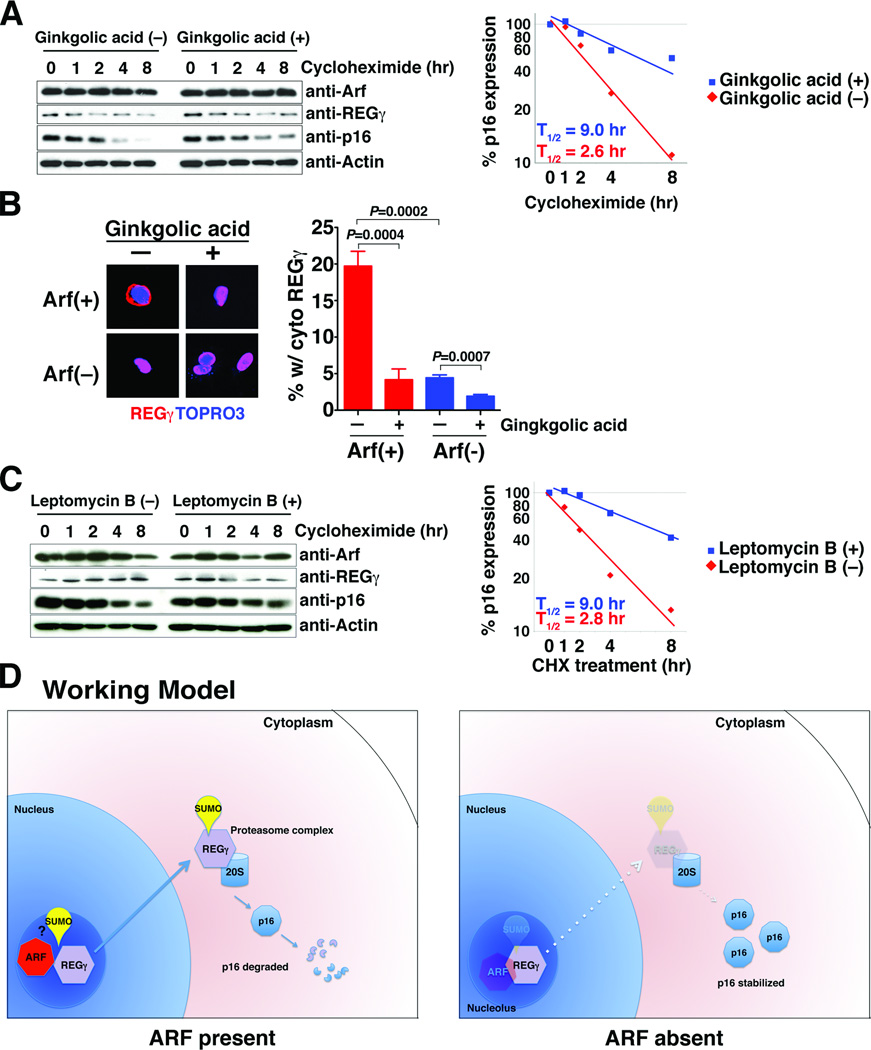
The cell-cycle regulatory gene INK4A-ARF (CDKN2A) has two alternative transcripts that produce entirely different proteins, namely p14(ARF) and p16, which have complementary functions as regulators of p53 and pRB tumor suppressor pathways, respectively. The unusual organization of INK4A-ARF has long led to speculation of a need for coordinated regulation of p14(ARF) and p16. We now show that p14(ARF) (ARF) regulates the stability of p16 protein in human cancer cell lines, as well as in mouse embryonic fibroblasts (MEFs). In particular, ARF promotes rapid degradation of p16 protein, which is mediated by the proteasome and, more specifically, by interaction of ARF with one of its subunits, REGγ. Furthermore, this ARF-dependent destabilization of p16 can be abrogated by knockdown of REGγ or by pharmacologic blockade of its nuclear export. Thus, our findings have uncovered a novel crosstalk of 2 key tumor suppressors mediated by a REGγ-dependent mechanism. The ability of ARF to control p16 stability may influence cell-cycle function.
Implications: The ability of ARF to control p16 stability may influence cell cycle function. Visual Overview: http://mcr.aacrjournals.org/content/current.
©2013 AACR.
Conflict of interest statement
Figures

References
-
- Ruas M, Peters G. The p16INK4a/CDKN2A tumor suppressor and its relatives. Biochim Biophys Acta. 1998;1378:F115–F177. - PubMed
-
- Sharpless NE. INK4a/ARF: a multifunctional tumor suppressor locus. Mutat Res. 2005;576:22–38. - PubMed
-
- Kamijo T, Zindy F, Roussel MF, Quelle DE, Downing JR, Ashmun RA, et al. Tumor suppression at the mouse INK4a locus mediated by the alternative reading frame product p19ARF. Cell. 1997;91:649–659. - PubMed
-
- Quelle DE, Zindy F, Ashmun RA, Sherr CJ. Alternative reading frames of the INK4a tumor suppressor gene encode two unrelated proteins capable of inducing cell cycle arrest. Cell. 1995;83:993–1000. - PubMed
-
- Sherr CJ. The INK4a/ARF network in tumour suppression. Nature reviews Molecular cell biology. 2001;2:731–737. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
