

Donor CD8+ T cells prevent Toxoplasma gondii de-encystation but fail to rescue the exhausted endogenous CD8+ T cell population
- PMID: 23817617
- PMCID: PMC3754203
- DOI: 10.1128/IAI.00784-12
Donor CD8+ T cells prevent Toxoplasma gondii de-encystation but fail to rescue the exhausted endogenous CD8+ T cell population
Abstract
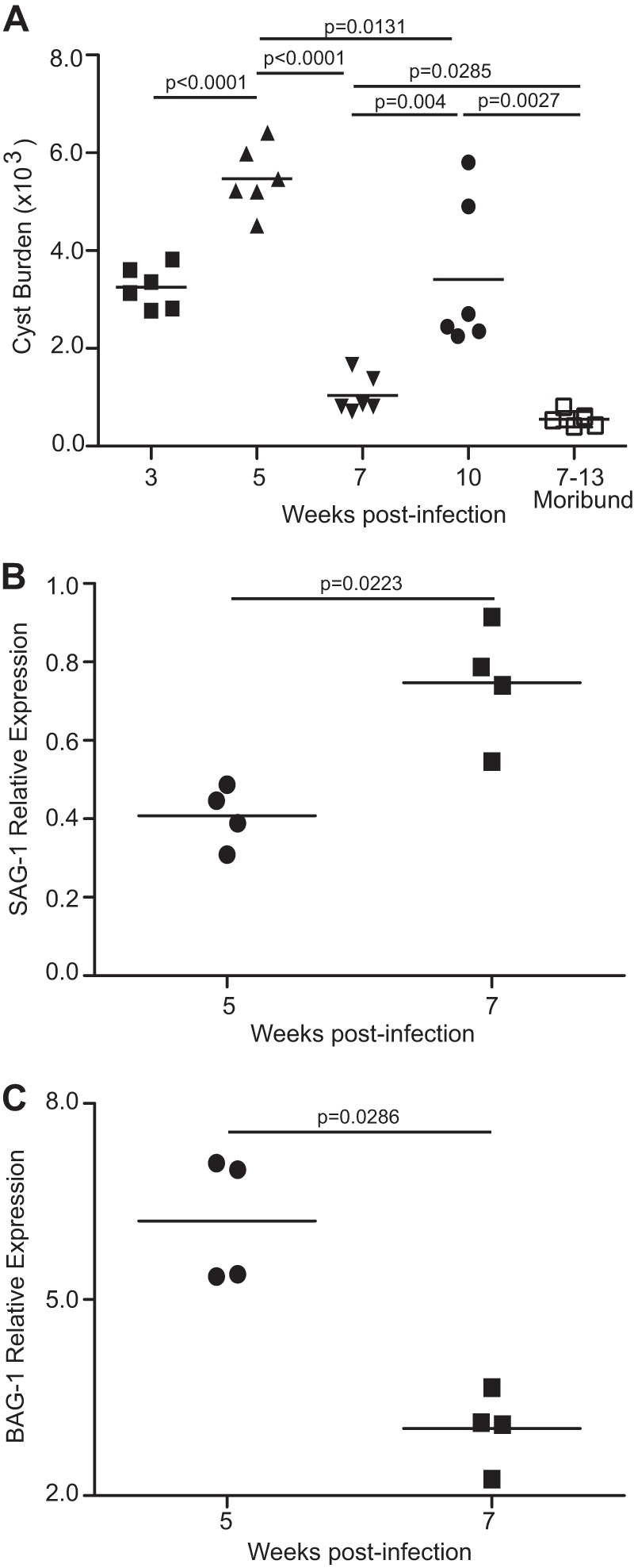
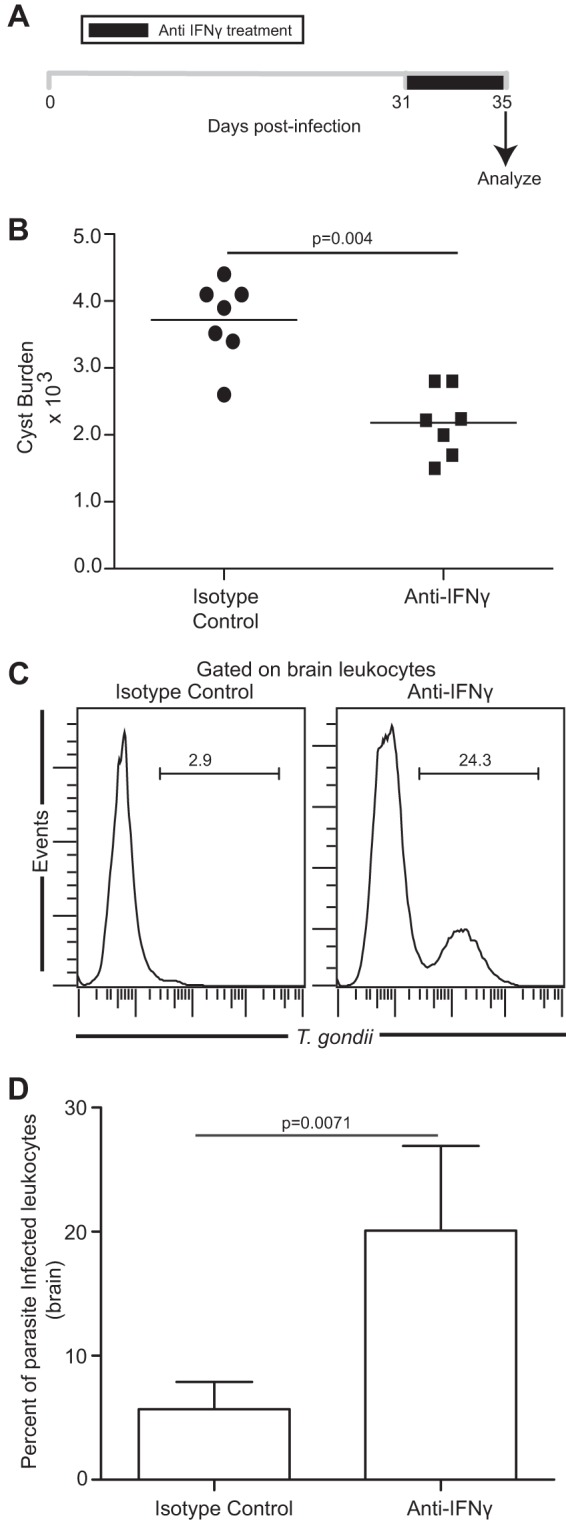
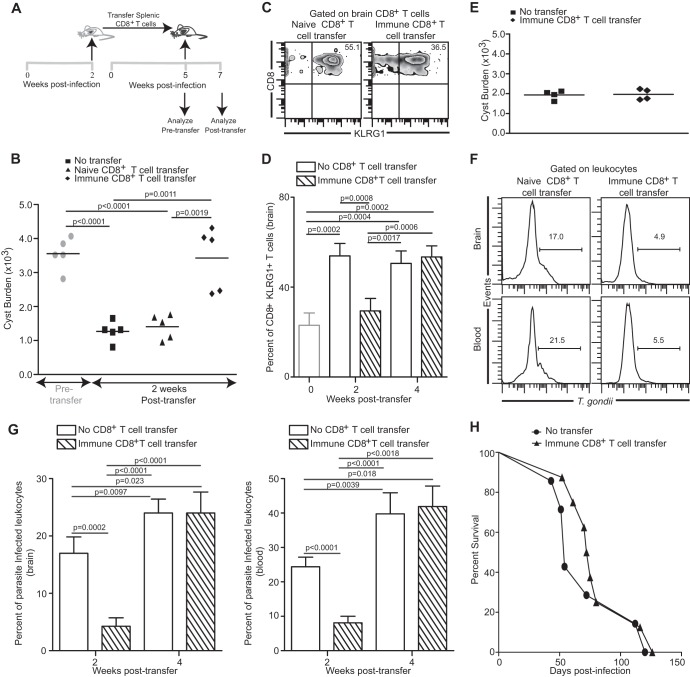
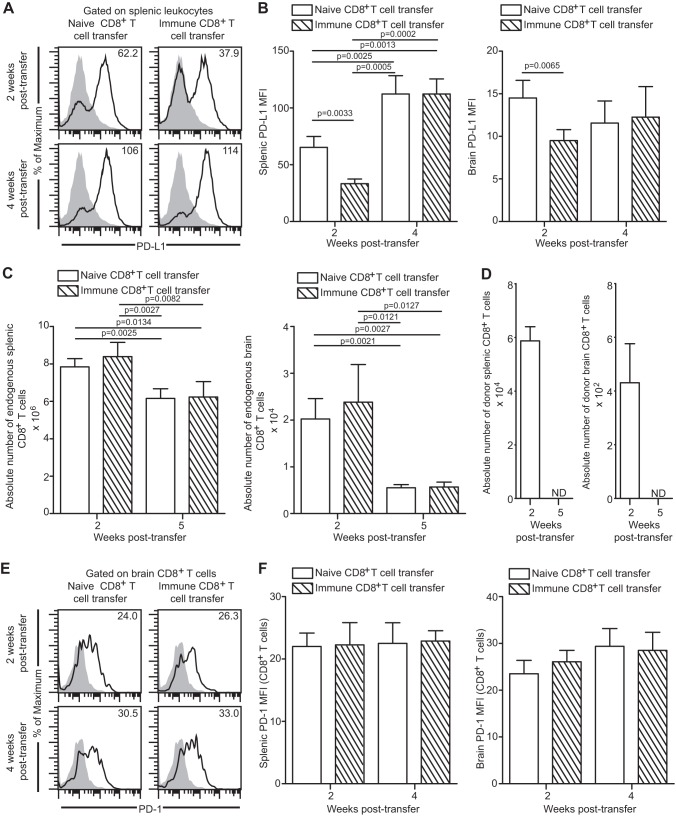
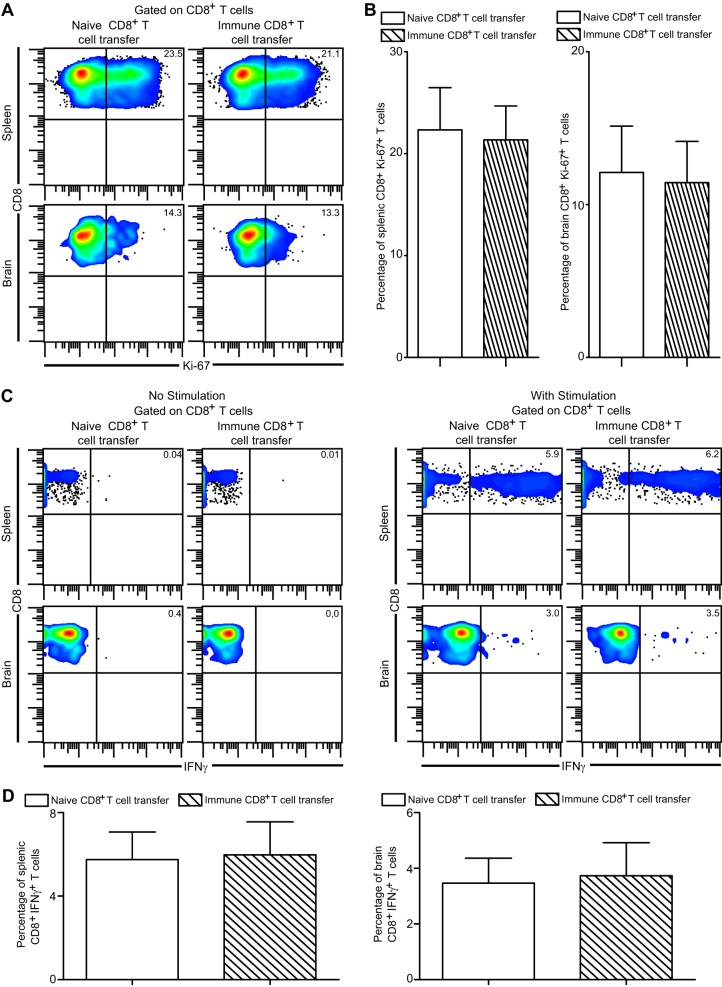
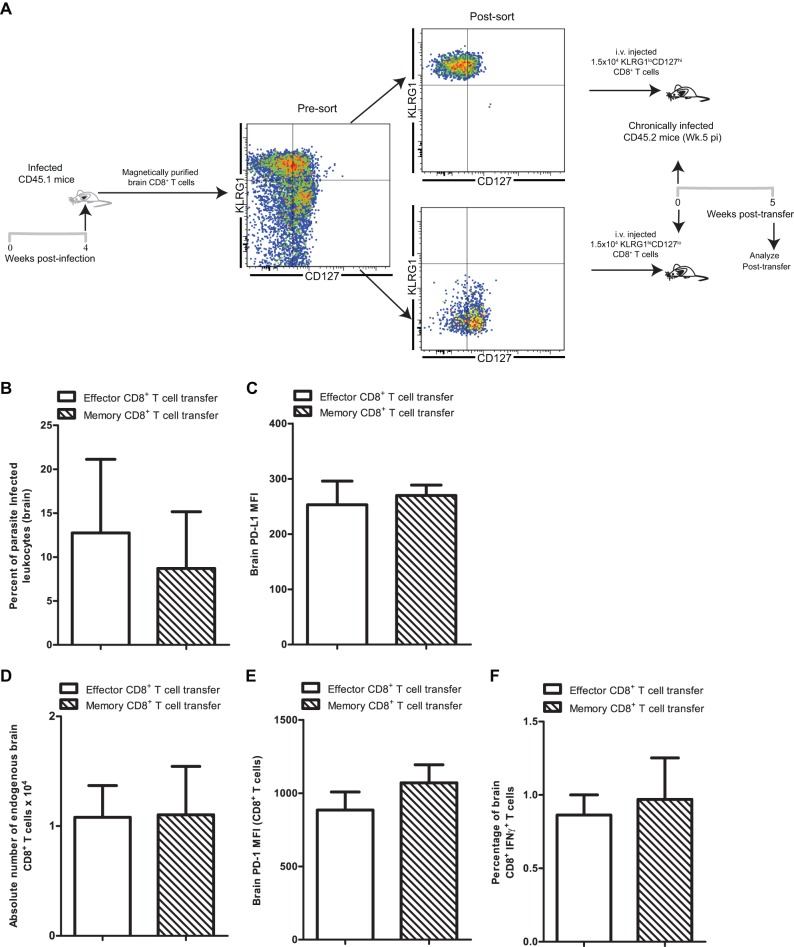
Functional exhaustion of CD8(+) T cells due to increased expression of inhibitory molecule PD-1 (Programmed Death-1) causes reactivation of latent disease during later phases of chronic toxoplasmosis. Onset of disease recrudescence results in decreased parasite cyst burden concomitant with parasites undergoing stage conversion from a primarily encysted, quiescent bradyzoite to a fast-replicating, highly motile tachyzoite. Thus, reduced cyst burden is one of the early hallmarks of disease recrudescence. This was further validated by depleting gamma interferon (IFN-γ), a cytokine known to control latent toxoplasmosis, in chronically infected prerecrudescent mice. Since CD8(+) T cells (an important source of IFN-γ) lose their functionality during the later phases of chronic toxoplasmosis, we next examined if adoptive transfer of functional CD8(+) T cells from acutely infected donors to the chronically infected prerecrudescent hosts could impede parasite de-encystation and rescue exhausted CD8(+) T cells. While the transfer of immune CD8(+) T cells temporarily restricted the breakdown of cysts, the exhausted endogenous CD8(+) T cell population was not rescued. Over time, the donor population got deleted, resulting in parasite de-encystation and host mortality. Considering that donor CD8(+) T cells fail to become long-lived, one of the cardinal features of memory CD8(+) T cells, it bears the implication that memory CD8 differentiation is impaired during chronic toxoplasmosis. Moreover, our data strongly suggest that while adoptive immunotherapy can prevent parasite de-encystation transiently, reduced antigen burden in the chronic phase by itself is insufficient for rescue of exhausted CD8(+) T cells. The conclusions of this study have profound ramifications in designing immunotherapeutics against chronic toxoplasmosis.
Figures
References
-
- Kur J, Holec-Gasior L, Hiszczynska-Sawicka E. 2009. Current status of toxoplasmosis vaccine development. Expert Rev. Vaccines 8:791–808 - PubMed
-
- Grant IH, Gold JW, Rosenblum M, Niedzwiecki D, Armstrong D. 1990. Toxoplasma gondii serology in HIV-infected patients: the development of central nervous system toxoplasmosis in AIDS. AIDS 4:519–521 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
