

Lifting della repression of Arabidopsis seed germination by nonproteolytic gibberellin signaling
- PMID: 23818171
- PMCID: PMC3729787
- DOI: 10.1104/pp.113.219451
Lifting della repression of Arabidopsis seed germination by nonproteolytic gibberellin signaling
Abstract
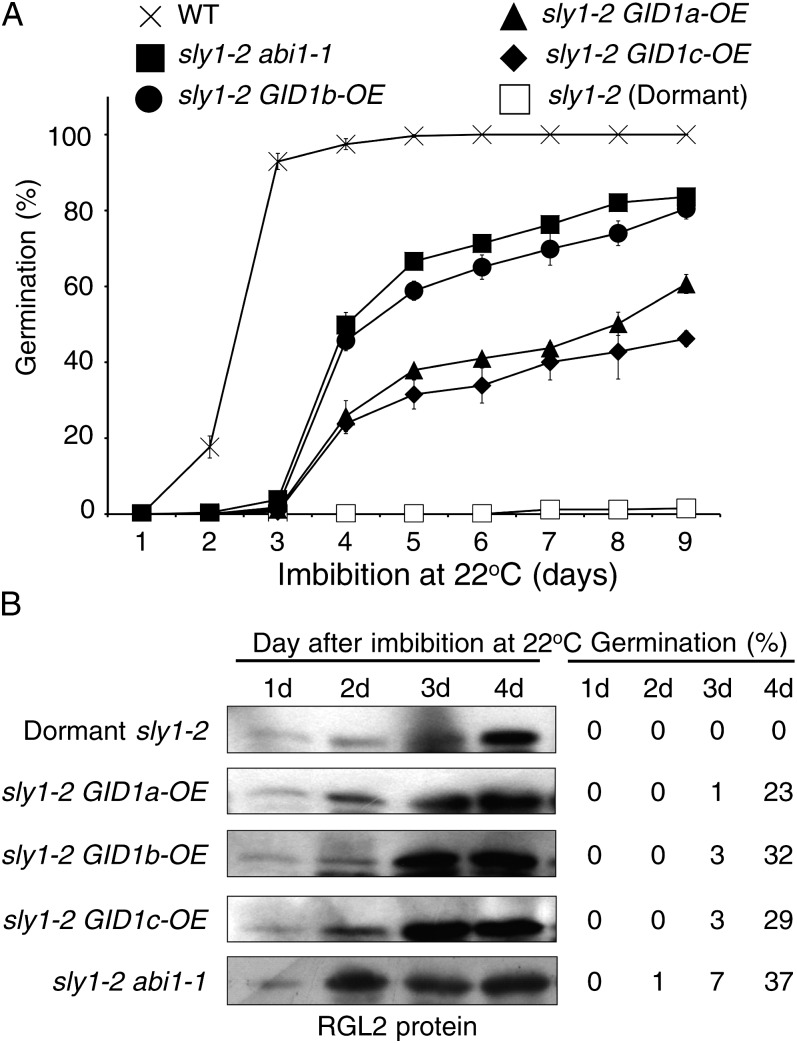
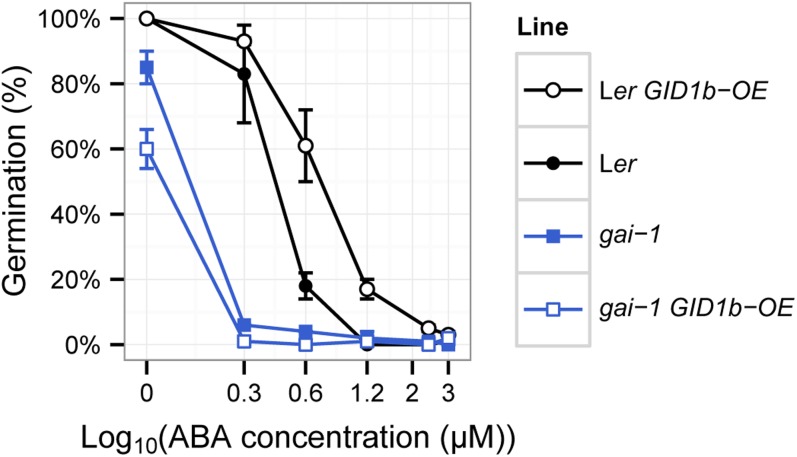
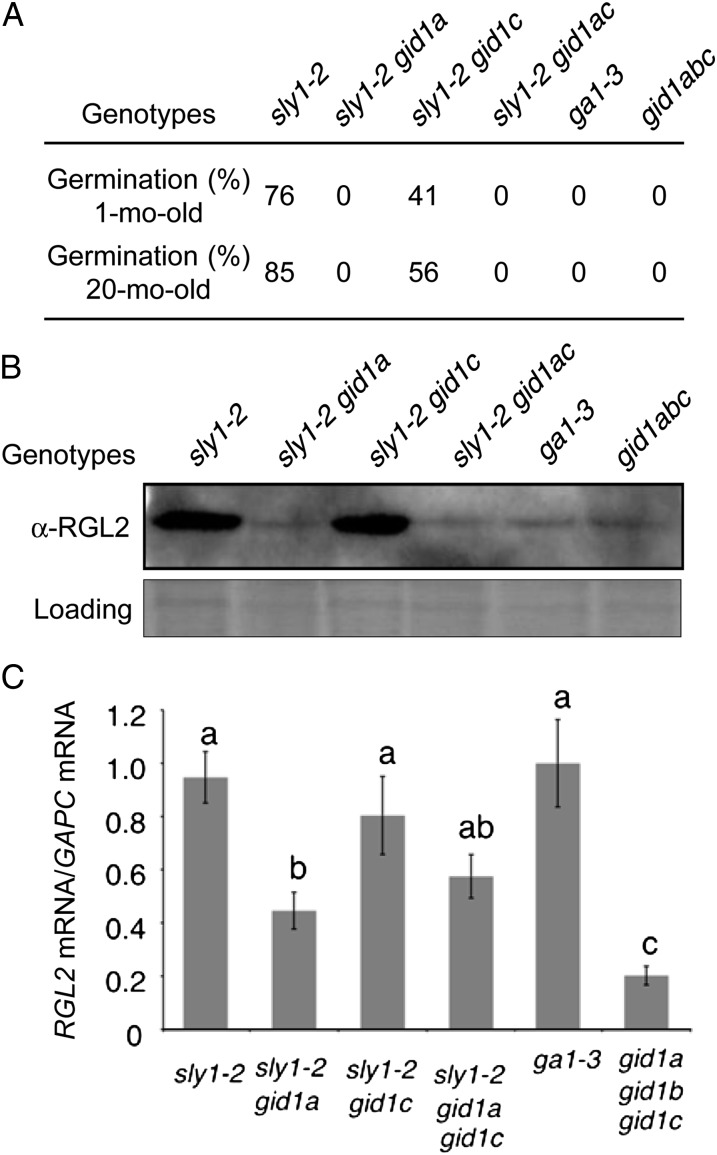
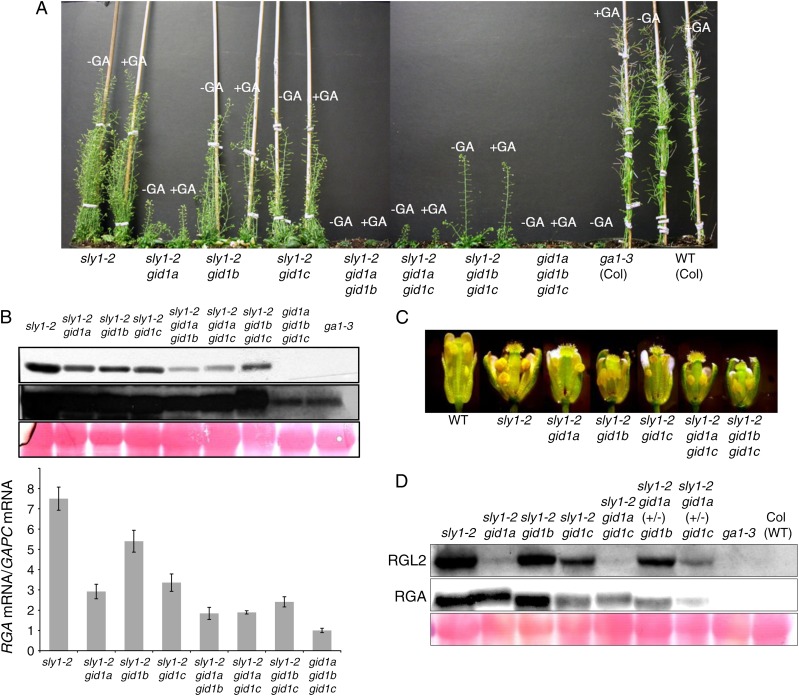
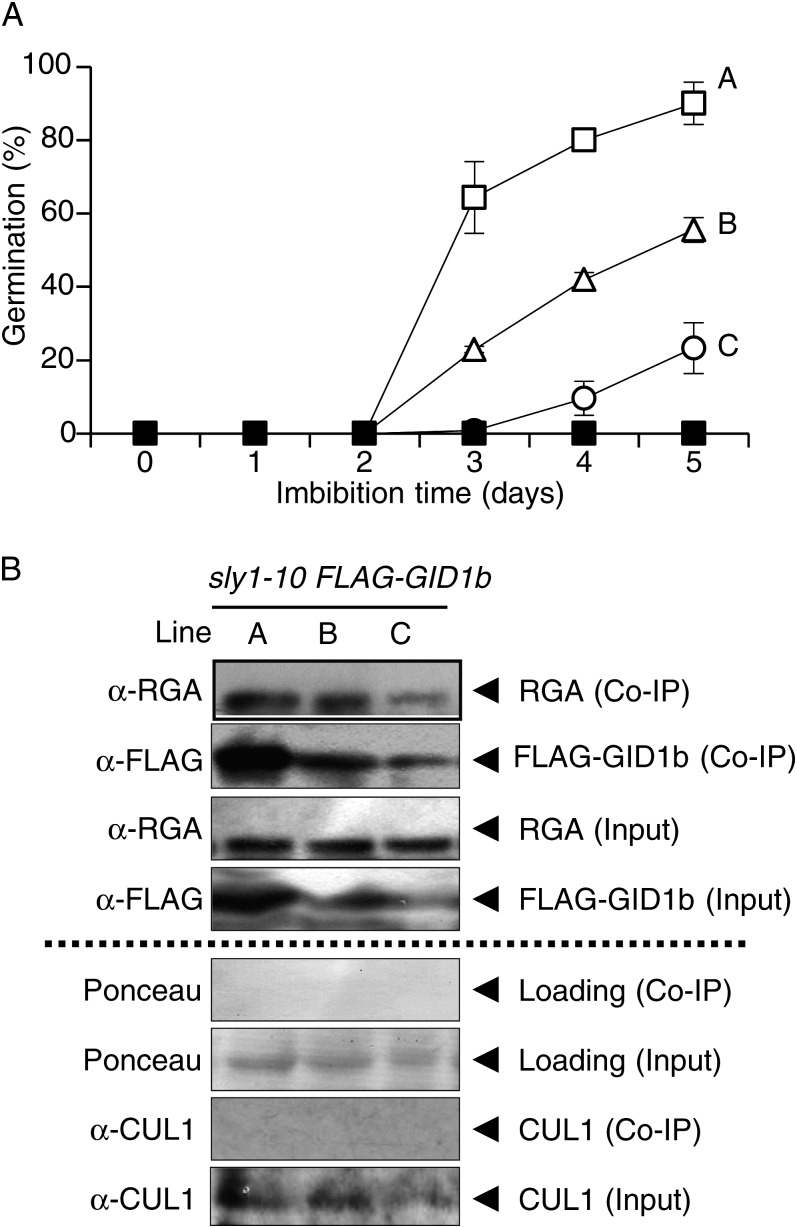
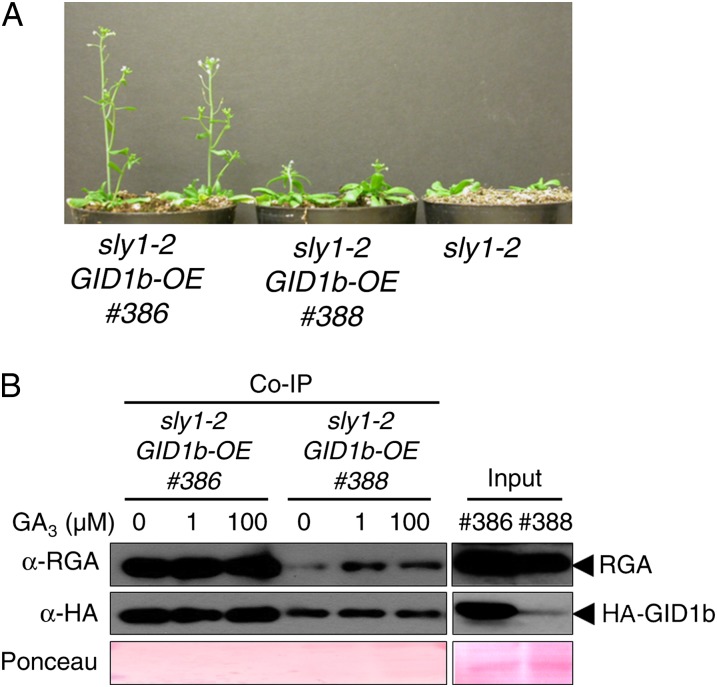
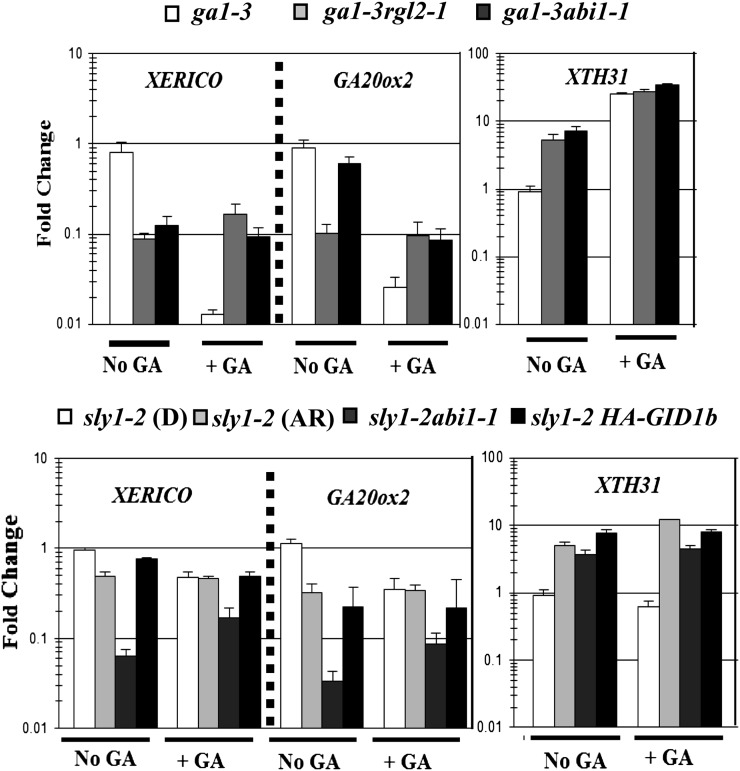
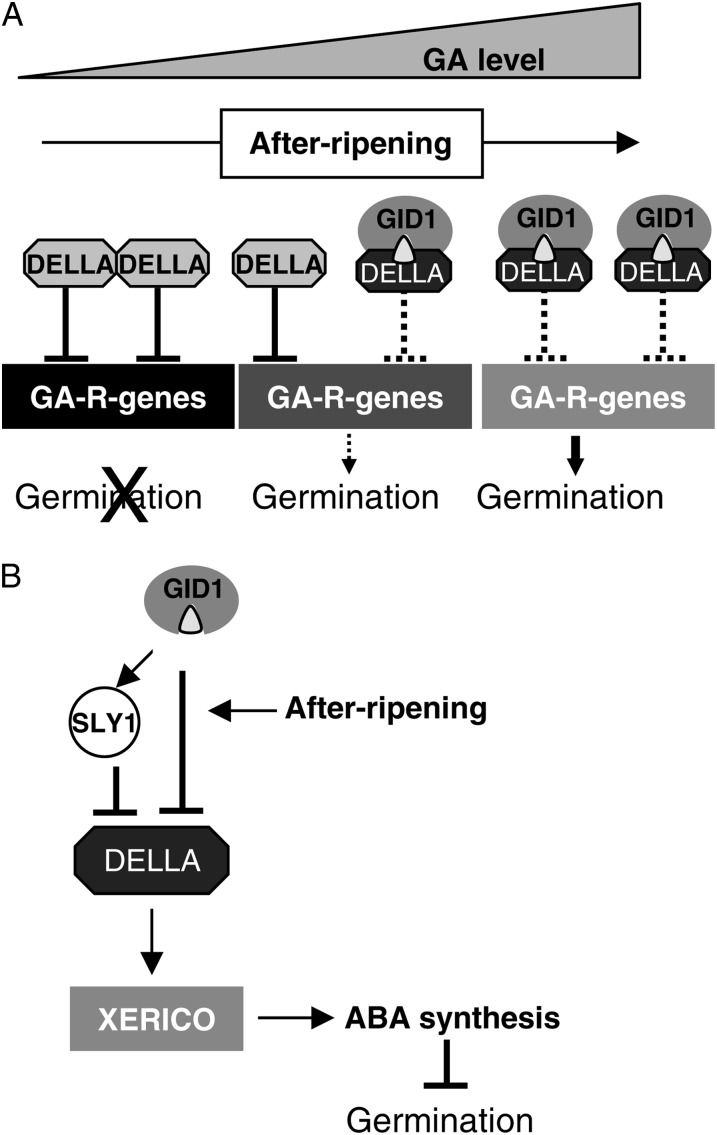
DELLA repression of Arabidopsis (Arabidopsis thaliana) seed germination can be lifted either through DELLA proteolysis by the ubiquitin-proteasome pathway or through proteolysis-independent gibberellin (GA) hormone signaling. GA binding to the GIBBERELLIN-INSENSITIVE DWARF1 (GID1) GA receptors stimulates GID1-GA-DELLA complex formation, which in turn triggers DELLA protein ubiquitination and proteolysis via the SCF(SLY1) E3 ubiquitin ligase and 26S proteasome. Although DELLA cannot be destroyed in the sleepy1-2 (sly1-2) F-box mutant, long dry after-ripening and GID1 overexpression can relieve the strong sly1-2 seed dormancy phenotype. It appears that sly1-2 seed dormancy results from abscisic acid (ABA) signaling downstream of DELLA, since dormant sly1-2 seeds accumulate high levels of ABA hormone and loss of ABA sensitivity rescues sly1-2 seed germination. DELLA positively regulates the expression of XERICO, an inducer of ABA biosynthesis. GID1b overexpression rescues sly1-2 germination through proteolysis-independent DELLA down-regulation associated with increased expression of GA-inducible genes and decreased ABA accumulation, apparently as a result of decreased XERICO messenger RNA levels. Higher levels of GID1 overexpression are associated with more efficient sly1 germination and increased GID1-GA-DELLA complex formation, suggesting that GID1 down-regulates DELLA through protein binding. After-ripening results in increased GA accumulation and GID1a-dependent GA signaling, suggesting that after-ripening triggers GA-stimulated GID1-GA-DELLA protein complex formation, which in turn blocks DELLA transcriptional activation of the XERICO inhibitor of seed germination.
Figures

References
-
- Bewley DJ, Black M (1994). Seeds: Physiology of Development and Germination. Plenum Press, New York
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
