

Inverse modulation of plant immune and brassinosteroid signaling pathways by the receptor-like cytoplasmic kinase BIK1
- PMID: 23818580
- PMCID: PMC3718091
- DOI: 10.1073/pnas.1302154110
Inverse modulation of plant immune and brassinosteroid signaling pathways by the receptor-like cytoplasmic kinase BIK1
Abstract
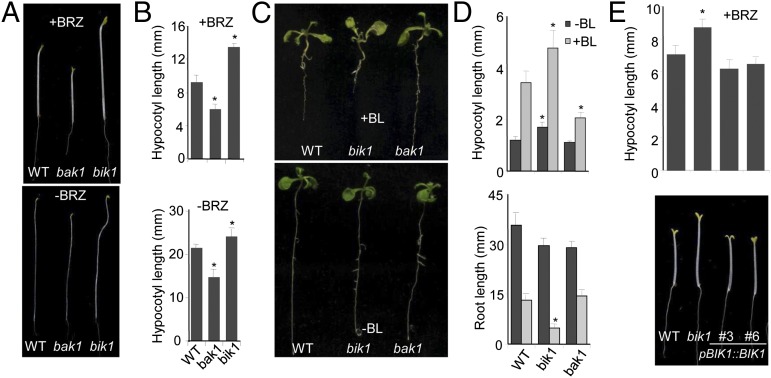
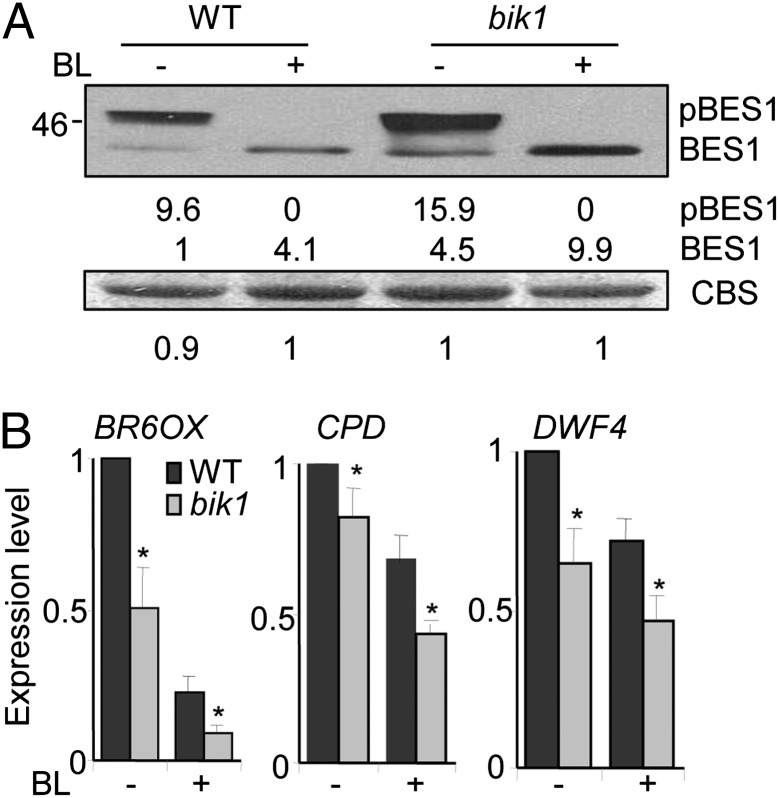
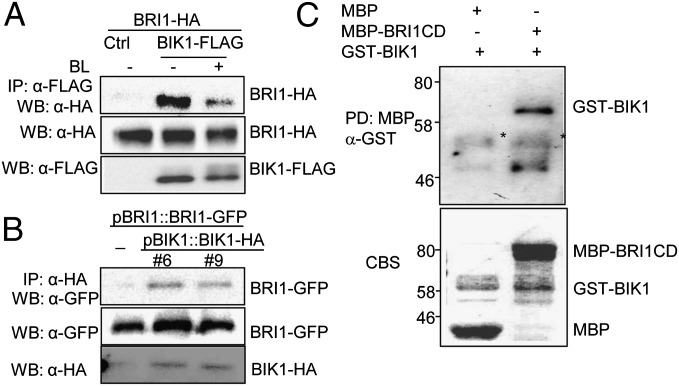
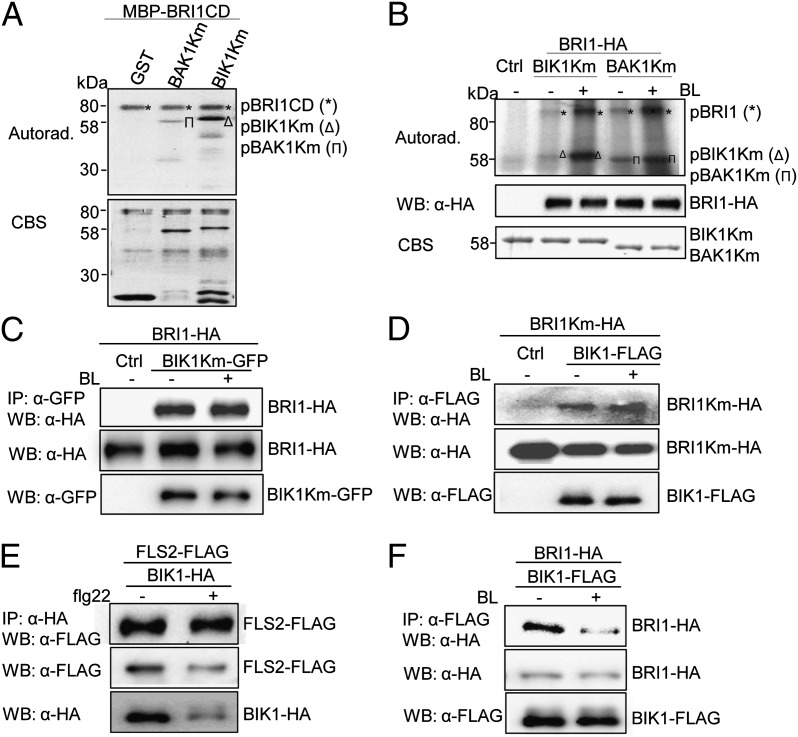
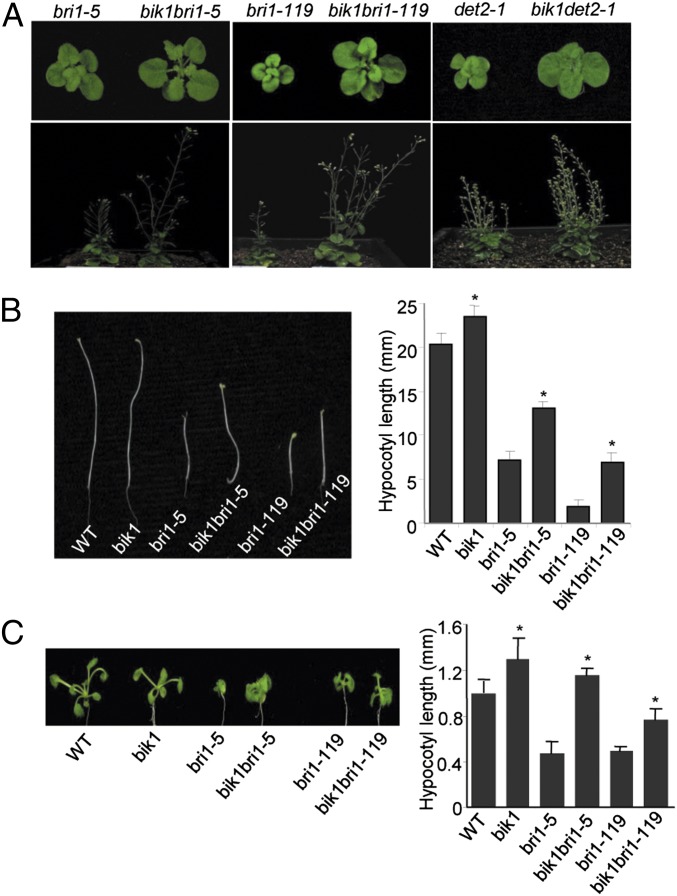
Maintaining active growth and effective immune responses is often costly for a living organism to survive. Fine-tuning the shared cross-regulators is crucial for metazoans and plants to make a trade-off between growth and immunity. The Arabidopsis regulatory receptor-like kinase BAK1 complexes with the receptor kinases FLS2 in bacterial flagellin-triggered immunity and BRI1 in brassinosteroid (BR)-mediated growth. BR homeostasis and signaling unidirectionally modulate FLS2-mediated immune responses at multiple levels. We have shown previously that BIK1, a receptor-like cytoplasmic kinase, is directly phosphorylated by BAK1 and associates with FLS2/BAK1 complex in transducing flagellin signaling. In contrast to its positive role in plant immunity, we report here that BIK1 acts as a negative regulator in BR signaling. The bik1 mutant displays various BR hypersensitive phenotypes accompanied with increased accumulation of de-phosphorylated BES1 proteins and transcriptional regulation of BZR1 and BES1 target genes. BIK1 associates with BRI1, and is released from BRI1 receptor upon BR treatment, which is reminiscent of FLS2-BIK1 complex dynamics in flagellin signaling. The ligand-induced release of BIK1 from receptor complexes is associated with BIK1 phosphorylation. However, in contrast to BAK1-dependent FLS2-BIK1 dissociation, BAK1 is dispensable for BRI1-BIK1 dissociation. Unlike FLS2 signaling which depends on BAK1 to phosphorylate BIK1, BRI1 directly phosphorylates BIK1 to transduce BR signaling. Thus, BIK1 relays the signaling in plant immunity and BR-mediated growth via distinct phosphorylation by BAK1 and BRI1, respectively. Our studies indicate that BIK1 mediates inverse functions in plant immunity and development via dynamic association with specific receptor complexes and differential phosphorylation events.
Keywords: BRI1-associated receptor kinase; botrytis-induced kinase 1; brassinosteroid insensitive 1; bri1-Ems-Suppressor 1; flagellin sensing 2.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Li J, Chory J. A putative leucine-rich repeat receptor kinase involved in brassinosteroid signal transduction. Cell. 1997;90(5):929–938. - PubMed
-
- Gómez-Gómez L, Boller T. FLS2: An LRR receptor-like kinase involved in the perception of the bacterial elicitor flagellin in Arabidopsis. Mol Cell. 2000;5(6):1003–1011. - PubMed
-
- Zipfel C, et al. Perception of the bacterial PAMP EF-Tu by the receptor EFR restricts Agrobacterium-mediated transformation. Cell. 2006;125(4):749–760. - PubMed
-
- Wang X, et al. Sequential transphosphorylation of the BRI1/BAK1 receptor kinase complex impacts early events in brassinosteroid signaling. Dev Cell. 2008;15(2):220–235. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
