

Diacylglycerol promotes centrosome polarization in T cells via reciprocal localization of dynein and myosin II
- PMID: 23818610
- PMCID: PMC3718145
- DOI: 10.1073/pnas.1306180110
Diacylglycerol promotes centrosome polarization in T cells via reciprocal localization of dynein and myosin II
Abstract
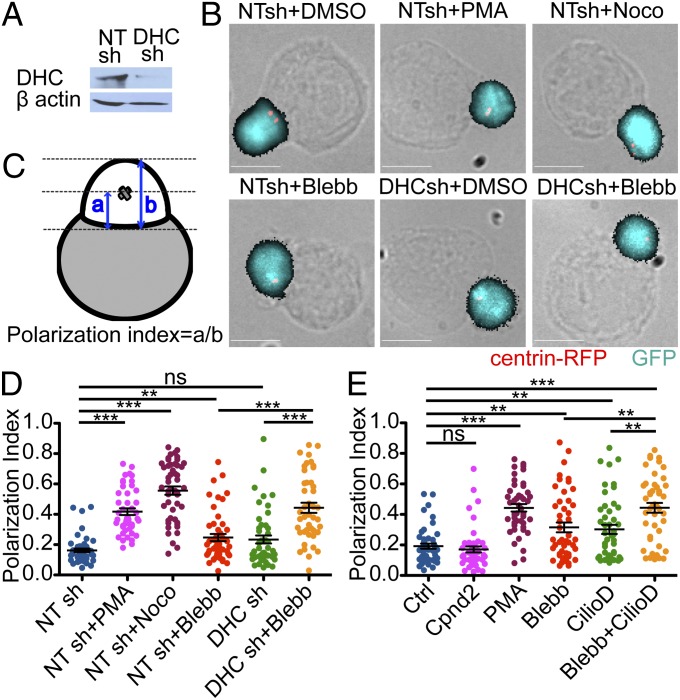
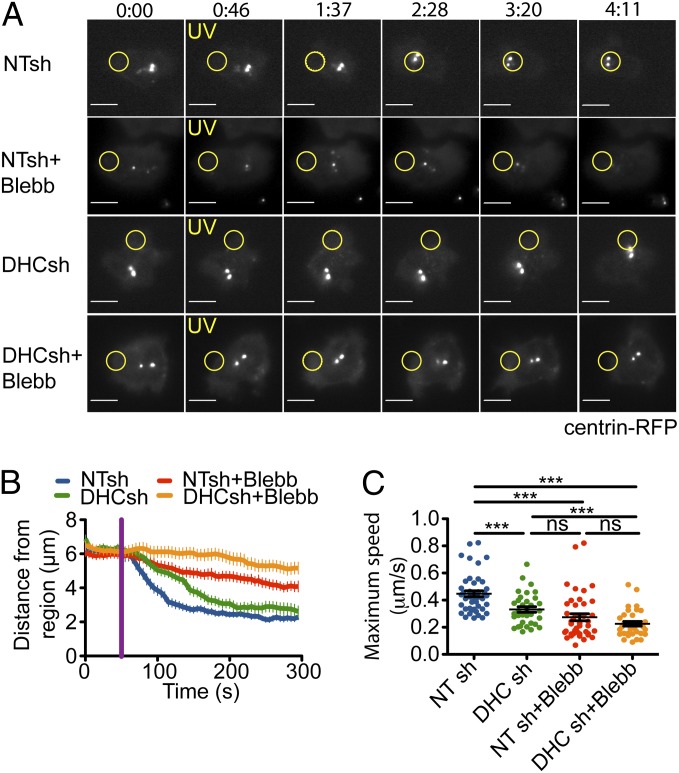
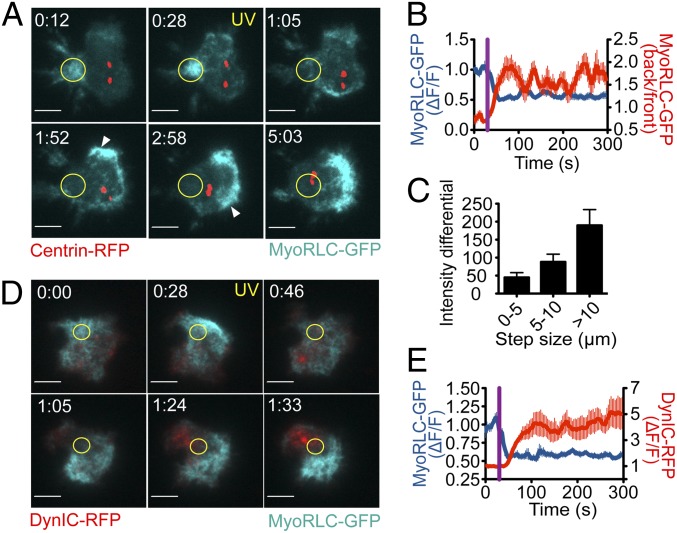
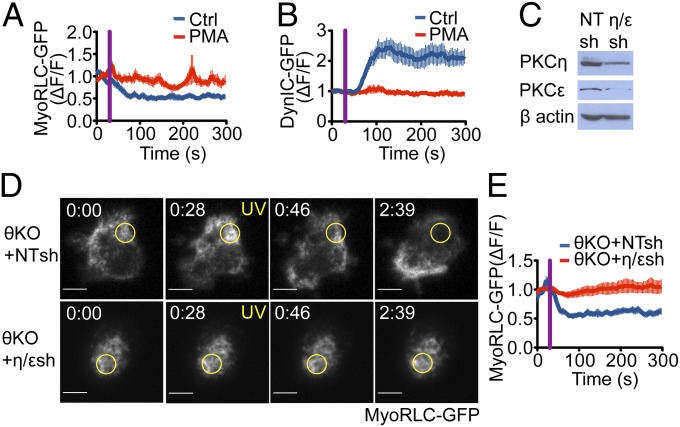
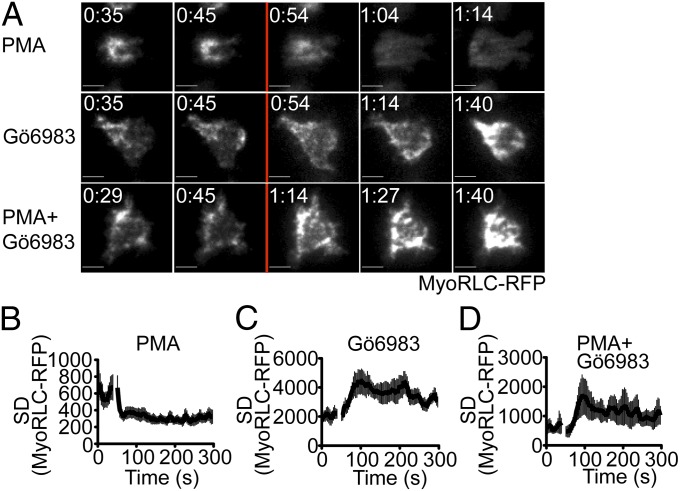
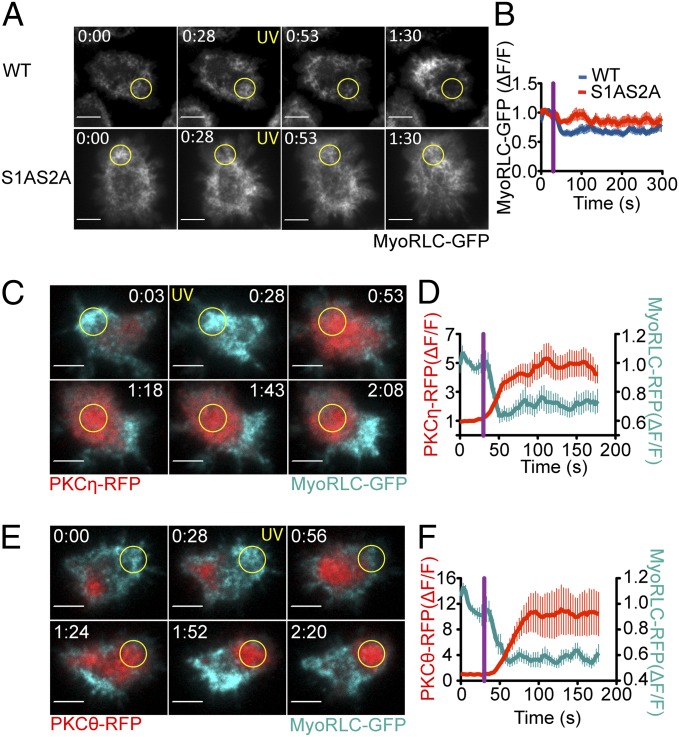
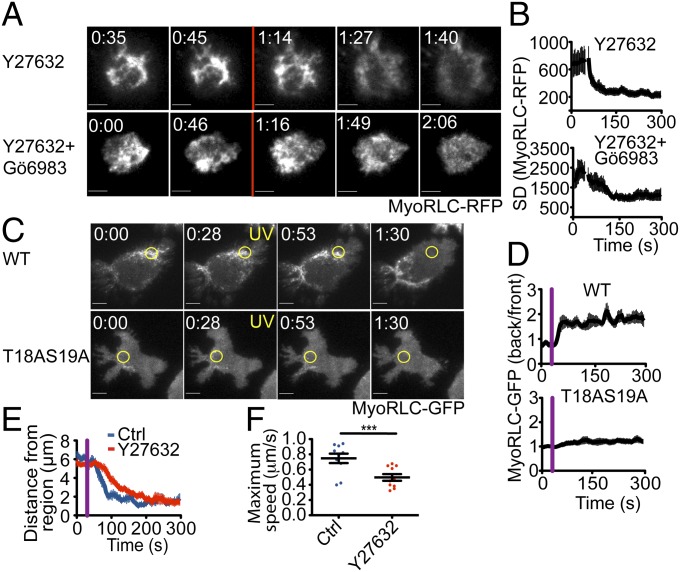
Centrosome reorientation to the immunological synapse maintains the specificity of T-cell effector function by facilitating the directional release of cytokines and cytolytic factors toward the antigen-presenting cell. This polarization response is driven by the localized accumulation of diacylglycerol, which recruits multiple protein kinase (PK)C isozymes to the synaptic membrane. Here, we used T-cell receptor (TCR) photoactivation and imaging methodology to demonstrate that PKCs control centrosome dynamics through the reciprocal localization of two motor complexes, dynein and nonmuscle myosin (NM)II. Dynein accumulated in the region of TCR stimulation, whereas NMII clustered in the back of the cell, behind the polarizing centrosome. PKC activity, which shaped both dynein and NMII accumulation within this framework, controlled NMII localization directly by phosphorylating inhibitory sites within the myosin regulatory light chain, thereby suppressing NMII clustering in the region of TCR stimulation. Concurrently, phosphorylation of distinct sites within myosin regulatory light chain by Rho kinase drove NMII clustering in areas behind the centrosome. These results reveal a role for NMII in T-cell polarity and demonstrate how it is regulated by upstream signals.
Keywords: cytoskeleton; microtubule-organizing center; polarity; signal transduction.
Figures
Similar articles
-
Actin clearance promotes polarized dynein accumulation at the immunological synapse.PLoS One. 2019 Jul 3;14(7):e0210377. doi: 10.1371/journal.pone.0210377. eCollection 2019. PLoS One. 2019. PMID: 31269031 Free PMC article.
-
From lipid second messengers to molecular motors: microtubule-organizing center reorientation in T cells.Immunol Rev. 2013 Nov;256(1):95-106. doi: 10.1111/imr.12116. Immunol Rev. 2013. PMID: 24117815 Free PMC article. Review.
-
Localized diacylglycerol drives the polarization of the microtubule-organizing center in T cells.Nat Immunol. 2009 Jun;10(6):627-35. doi: 10.1038/ni.1734. Nat Immunol. 2009. PMID: 19430478
-
A cascade of protein kinase C isozymes promotes cytoskeletal polarization in T cells.Nat Immunol. 2011 May 22;12(7):647-54. doi: 10.1038/ni.2033. Nat Immunol. 2011. PMID: 21602810 Free PMC article.
-
The role of the cytoskeleton at the immunological synapse.Immunol Rev. 2013 Nov;256(1):107-17. doi: 10.1111/imr.12117. Immunol Rev. 2013. PMID: 24117816 Free PMC article. Review.
Cited by
-
Regulation of T cell signalling by membrane lipids.Nat Rev Immunol. 2016 Nov;16(11):690-701. doi: 10.1038/nri.2016.103. Epub 2016 Oct 10. Nat Rev Immunol. 2016. PMID: 27721483 Review.
-
Cytotoxic T Lymphocyte Activation Signals Modulate Cytoskeletal Dynamics and Mechanical Force Generation.Front Immunol. 2022 Mar 16;13:779888. doi: 10.3389/fimmu.2022.779888. eCollection 2022. Front Immunol. 2022. PMID: 35371019 Free PMC article.
-
Dynamic microtubules regulate cellular contractility during T-cell activation.Proc Natl Acad Sci U S A. 2017 May 23;114(21):E4175-E4183. doi: 10.1073/pnas.1614291114. Epub 2017 May 10. Proc Natl Acad Sci U S A. 2017. PMID: 28490501 Free PMC article.
-
Inducible Polarized Secretion of Exosomes in T and B Lymphocytes.Int J Mol Sci. 2020 Apr 10;21(7):2631. doi: 10.3390/ijms21072631. Int J Mol Sci. 2020. PMID: 32290050 Free PMC article. Review.
-
Diacylglycerol Kinases: Shaping Diacylglycerol and Phosphatidic Acid Gradients to Control Cell Polarity.Front Cell Dev Biol. 2016 Nov 29;4:140. doi: 10.3389/fcell.2016.00140. eCollection 2016. Front Cell Dev Biol. 2016. PMID: 27965956 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
