

Activation of influenza viruses by proteases from host cells and bacteria in the human airway epithelium
- PMID: 23821437
- PMCID: PMC7108517
- DOI: 10.1111/2049-632X.12053
Activation of influenza viruses by proteases from host cells and bacteria in the human airway epithelium
Abstract
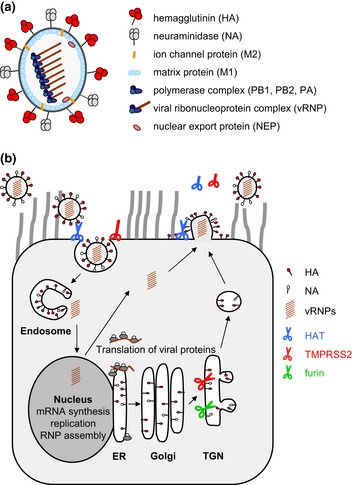
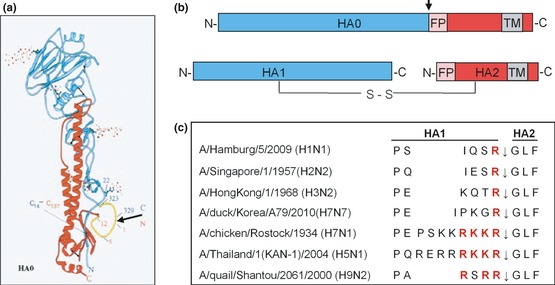
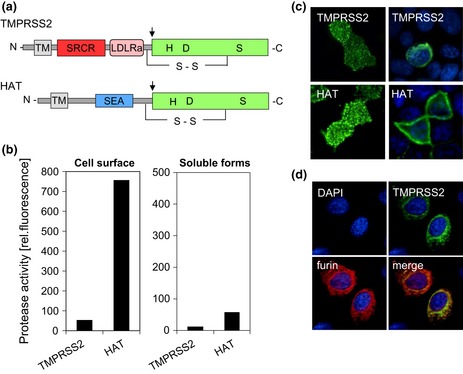
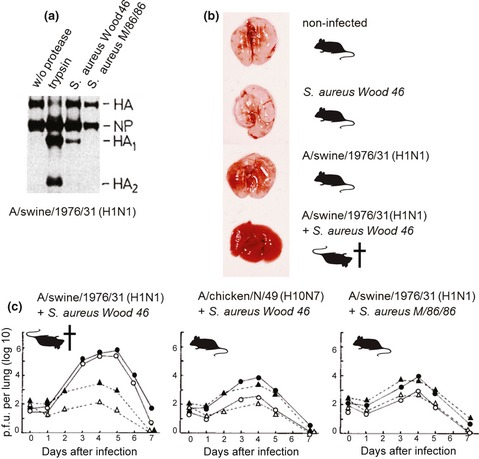
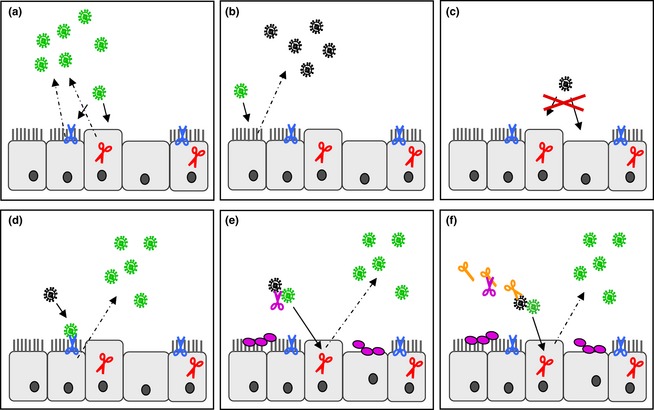
Influenza is an acute infection of the respiratory tract, which affects each year millions of people. Influenza virus infection is initiated by the surface glycoprotein hemagglutinin (HA) through receptor binding and fusion of viral and endosomal membranes. HA is synthesized as a precursor protein and requires cleavage by host cell proteases to gain its fusion capacity. Although cleavage of HA is crucial for virus infectivity, little was known about relevant proteases in the human airways for a long time. Recent progress in the identification and characterization of HA-activating host cell proteases has been considerable however and supports the idea of targeting HA cleavage as a novel approach for influenza treatment. Interestingly, certain bacteria have been demonstrated to support HA activation either by secreting proteases that cleave HA or due to activation of cellular proteases and thereby may contribute to virus spread and enhanced pathogenicity. In this review, we give an overview on activation of influenza viruses by proteases from host cells and bacteria with the main focus on recent progress on HA cleavage by proteases HAT and TMPRSS2 in the human airway epithelium. In addition, we outline investigations of HA-activating proteases as potential drug targets for influenza treatment.
Keywords: HAT; TMPRSS2; influenza treatment by protease inhibitors; influenza virus hemagglutinin; proteolytic cleavage; viral-bacterial pneumonia.
© 2013 Federation of European Microbiological Societies. Published by John Wiley & Sons Ltd. All rights reserved.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Afar DE, Vivanco I, Hubert RS, Kuo J, Chen E, Saffran DC, Raitano AB & Jakobovits A (2001) Catalytic cleavage of the androgen‐regulated TMPRSS2 protease results in its secretion by prostate and prostate cancer epithelia. Cancer Res 61: 1686–1692. - PubMed
-
- Bertram S, Heurich A, Lavender H, Gierer S, Danisch S, Perin P, Lucas JM, Nelson PS, Pöhlmann S & Soilleux EJ (2012) Influenza and SARS‐coronavirus activating proteases TMPRSS2 and HAT are expressed at multiple sites in human respiratory and gastrointestinal tracts. PLoS ONE 7: e35876. - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
