

Cloning and characterization of a critical regulator for preharvest sprouting in wheat
- PMID: 23821595
- PMCID: PMC3761307
- DOI: 10.1534/genetics.113.152330
Cloning and characterization of a critical regulator for preharvest sprouting in wheat
Abstract
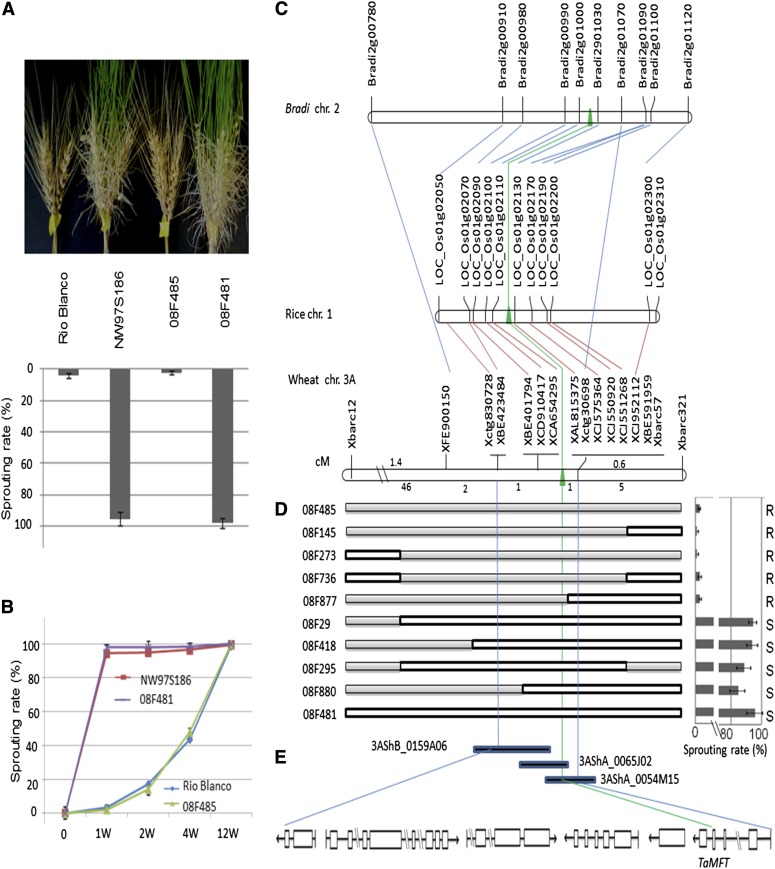
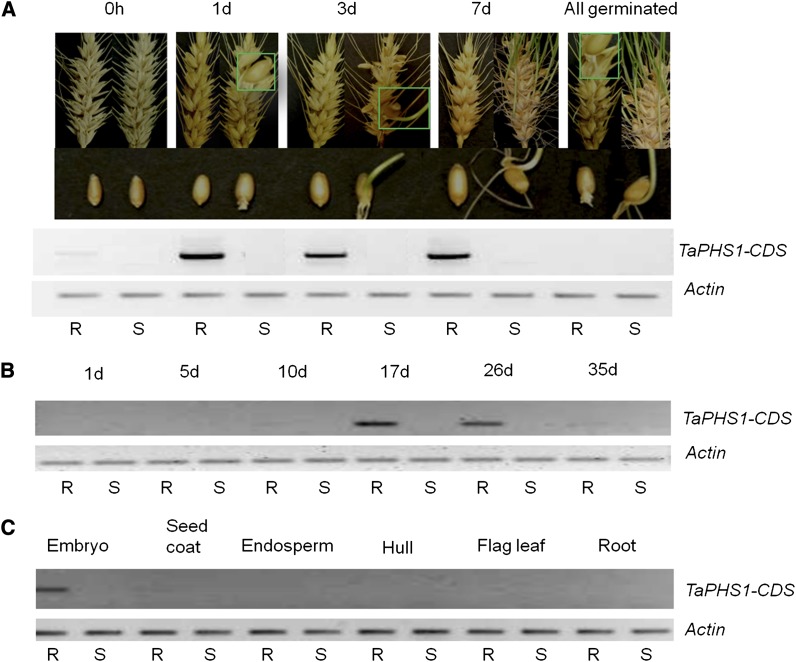
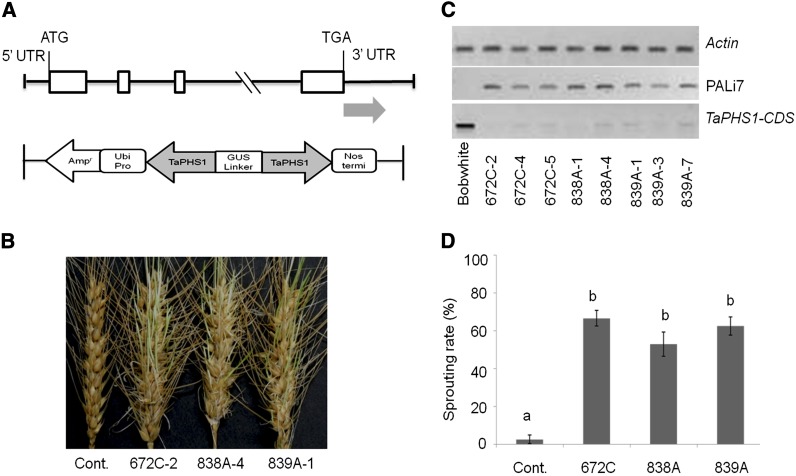
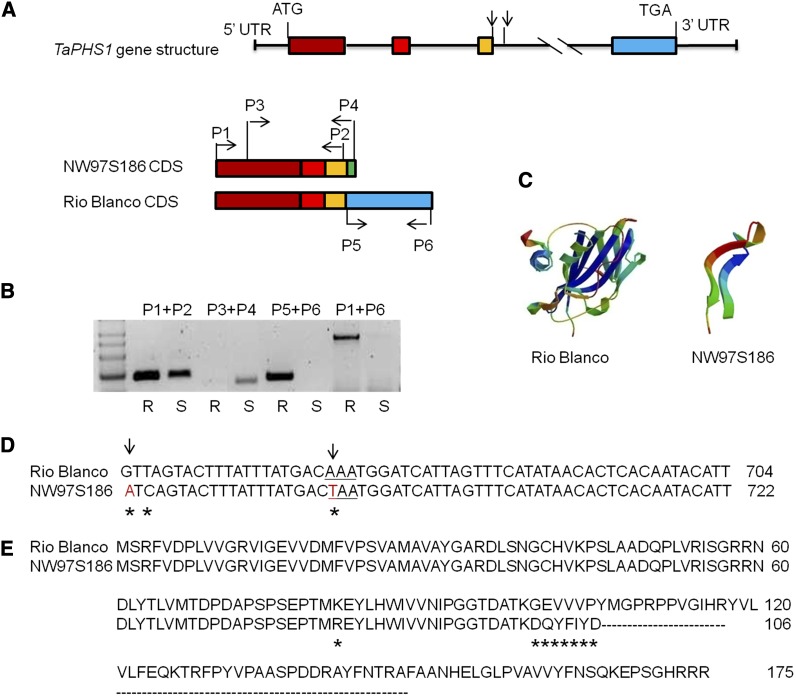
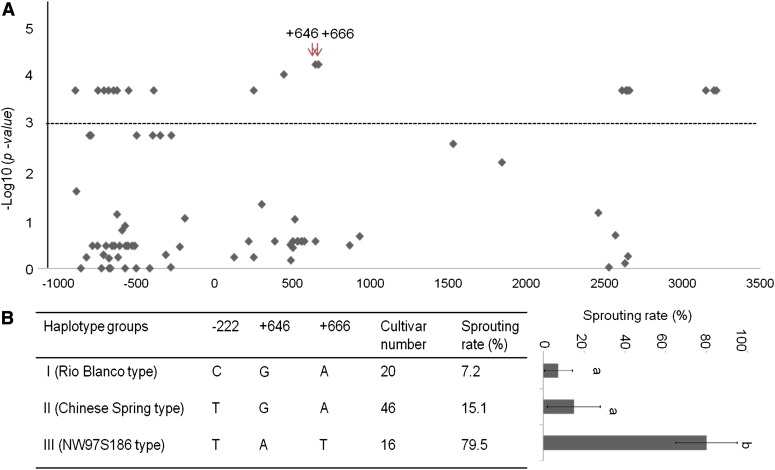
Sprouting of grains in mature spikes before harvest is a major problem in wheat (Triticum aestivum) production worldwide. We cloned and characterized a gene underlying a wheat quantitative trait locus (QTL) on the short arm of chromosome 3A for preharvest sprouting (PHS) resistance in white wheat using comparative mapping and map-based cloning. This gene, designated TaPHS1, is a wheat homolog of a MOTHER OF FLOWERING TIME (TaMFT)-like gene. RNA interference-mediated knockdown of the gene confirmed that TaPHS1 positively regulates PHS resistance. We discovered two causal mutations in TaPHS1 that jointly altered PHS resistance in wheat. One GT-to-AT mutation generates a mis-splicing site, and the other A-to-T mutation creates a premature stop codon that results in a truncated nonfunctional transcript. Association analysis of a set of wheat cultivars validated the role of the two mutations on PHS resistance. The molecular characterization of TaPHS1 is significant for expediting breeding for PHS resistance to protect grain yield and quality in wheat production.
Keywords: DNA marker; Preharvesting sprouting; RNA interference; gene clone; wheat abiotic stress.
Figures
References
-
- Altpeter F., Vasil V., Srivastava V., Stoger E., Vasil I. K., 1996. Accelerated production of transgenic wheat (Triticum aestivum L.) plants. Plant Cell Rep. 16: 12–17. - PubMed
-
- Baumlein H., Nagy I., Villarroel R., Inze D., Wobus U., 1992. Cis-analysis of a seed protein gene promoter: The conservative RY repeat CATGCATG within the legumin box is essential for tissue-specific expression of a legumin gene. Plant J. 2: 233–239. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
