

Dynamics of nucleoid structure regulated by mitochondrial fission contributes to cristae reformation and release of cytochrome c
- PMID: 23821750
- PMCID: PMC3718159
- DOI: 10.1073/pnas.1301951110
Dynamics of nucleoid structure regulated by mitochondrial fission contributes to cristae reformation and release of cytochrome c
Abstract
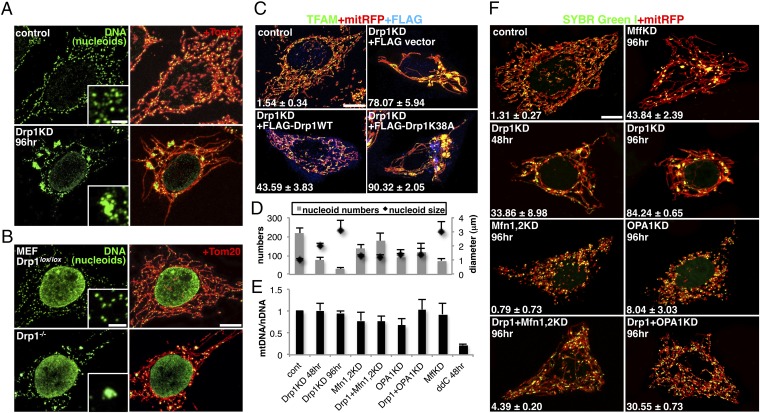
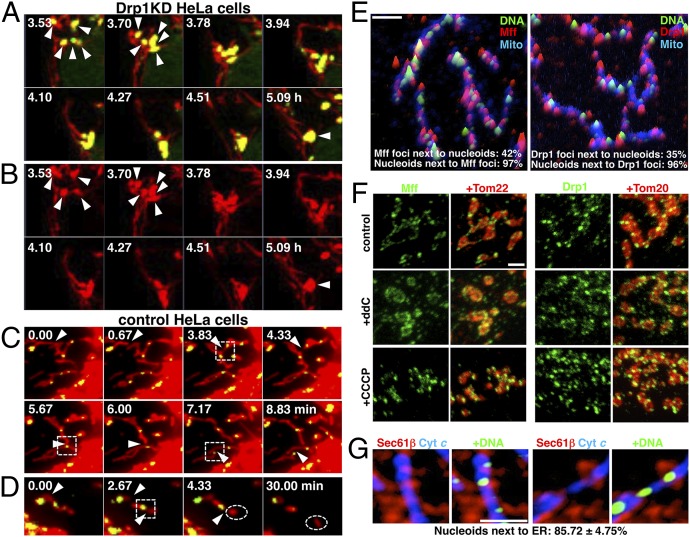
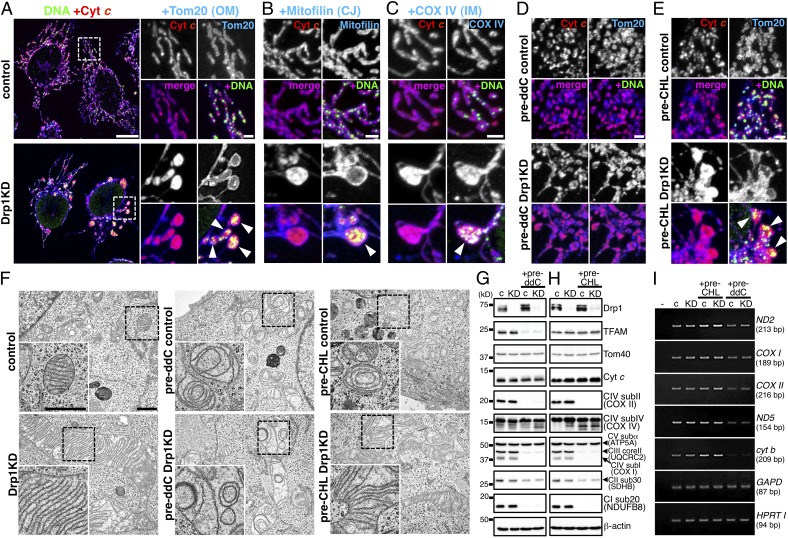
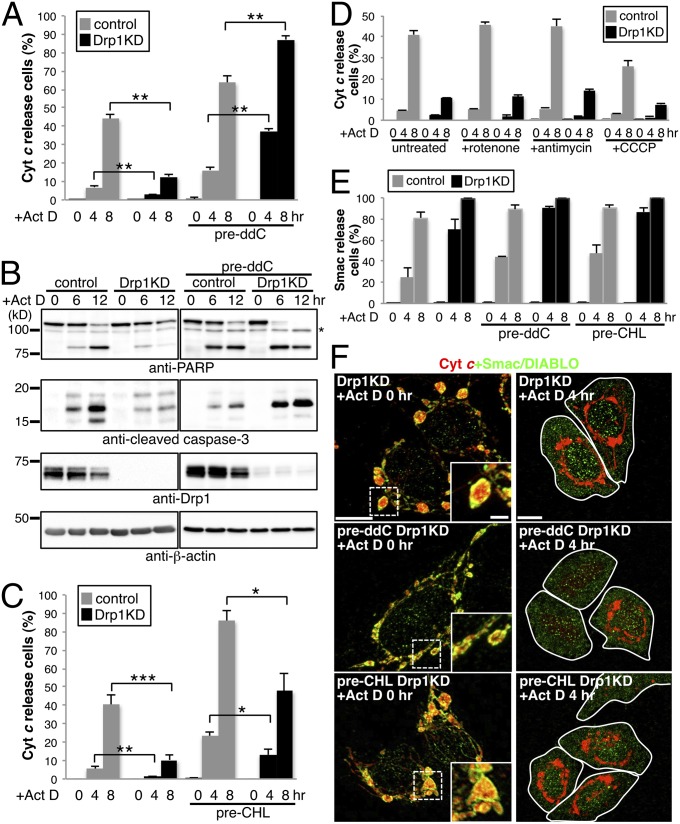
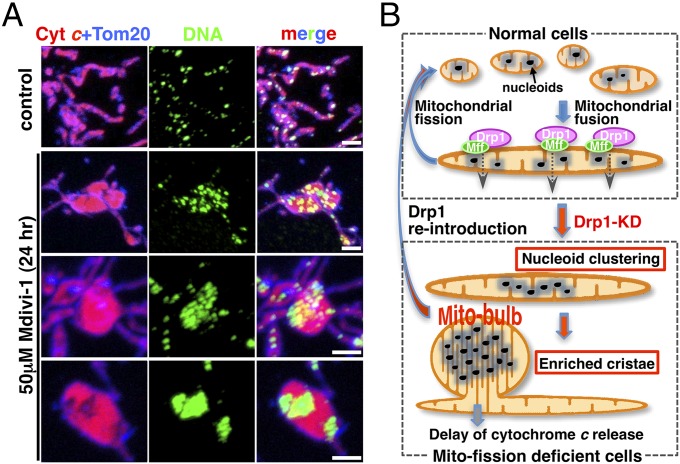
Mammalian cells typically contain thousands of copies of mitochondrial DNA assembled into hundreds of nucleoids. Here we analyzed the dynamic features of nucleoids in terms of mitochondrial membrane dynamics involving balanced fusion and fission. In mitochondrial fission GTPase dynamin-related protein (Drp1)-deficient cells, nucleoids were enlarged by their clustering within hyperfused mitochondria. In normal cells, mitochondrial fission often occurred adjacent to nucleoids, since localization of Mff and Drp1 is dependent on the nucleoids. Thus, mitochondrial fission adjacent to nucleoids should prevent their clustering by maintaining small and fragmented nucleoids. The enhanced clustering of nucleoids resulted in the formation of highly stacked cristae structures in enlarged bulb-like mitochondria (mito-bulbs). Enclosure of proapoptotic factor cytochrome c, but not of Smac/DIABLO, into the highly stacked cristae suppressed its release from mitochondria under apoptotic stimuli. In the absence of nucleoids, Drp1 deficiency failed to form mito-bulbs and to protect against apoptosis. Thus, mitochondrial dynamics by fission and fusion play a critical role in controlling mitochondrial nucleoid structures, contributing to cristae reformation and the proapoptotic status of mitochondria.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
