

Tunable signal processing through a kinase control cycle: the IKK signaling node
- PMID: 23823243
- PMCID: PMC3699752
- DOI: 10.1016/j.bpj.2013.05.013
Tunable signal processing through a kinase control cycle: the IKK signaling node
Abstract
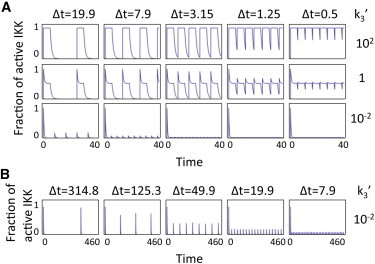
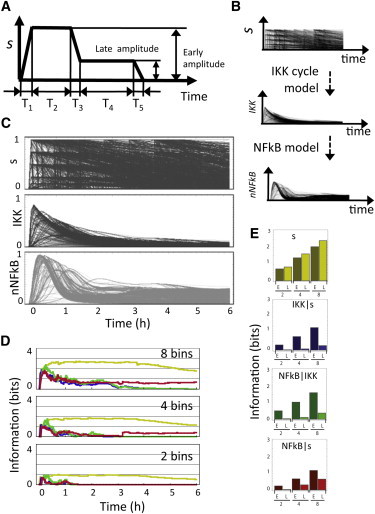
The transcription factor NFκB, a key component of the immune system, shows intricate stimulus-specific temporal dynamics. Those dynamics are thought to play a role in controlling the physiological response to cytokines and pathogens. Biochemical evidence suggests that the NFκB inducing kinase, IKK, a signaling hub onto which many signaling pathways converge, is regulated via a regulatory cycle comprising a poised, an active, and an inactive state. We hypothesize that it operates as a modulator of signal dynamics, actively reshaping the signals generated at the receptor proximal level. Here we show that a regulatory cycle can function in at least three dynamical regimes, tunable by regulating a single kinetic parameter. In particular, the simplest three-state regulatory cycle can generate signals with two well-defined phases, each with distinct coding capabilities in terms of the information they can carry about the stimulus. We also demonstrate that such a kinase cycle can function as a signal categorizer classifying diverse incoming signals into outputs with a limited set of temporal activity profiles. Finally, we discuss the extension of the results to other regulatory motifs that could be understood in terms of the regimes of the three-state cycle.
Copyright © 2013 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Figures
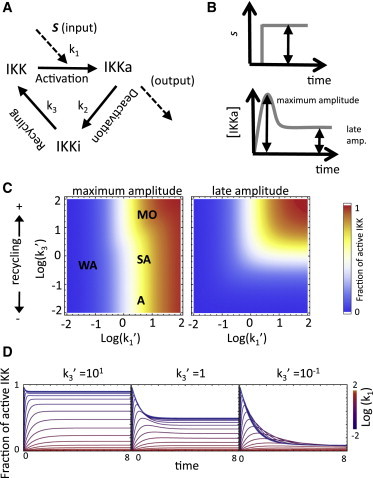
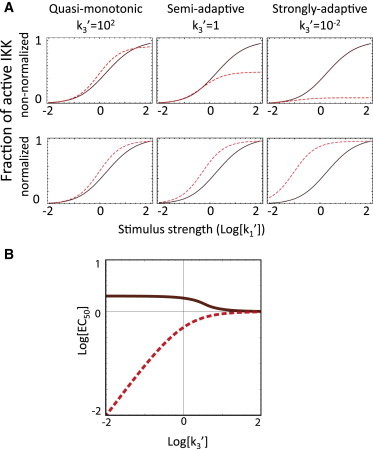
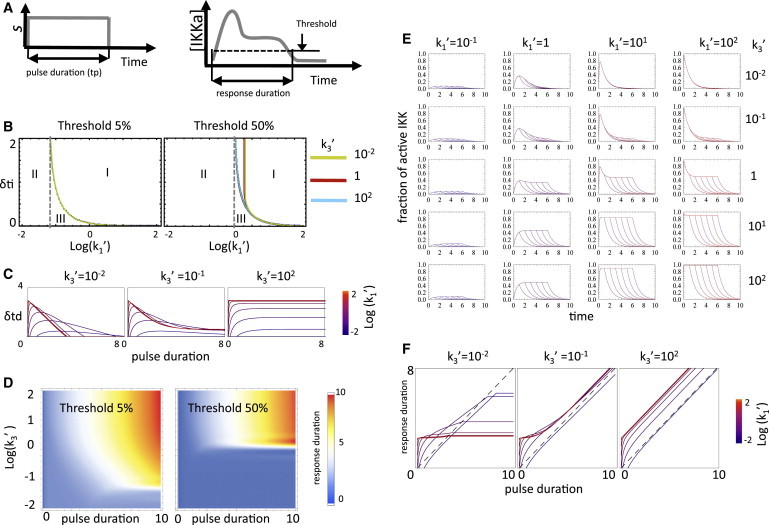
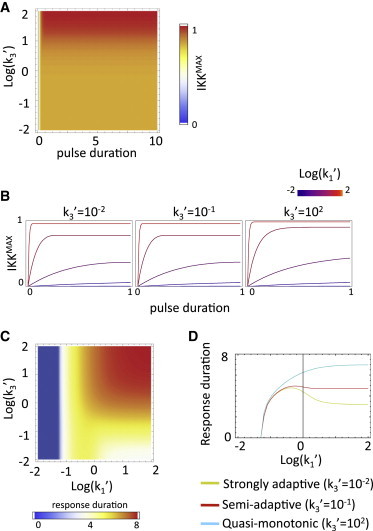
References
-
- Hayden M.S., Ghosh S. Shared principles in NF-κB signaling. Cell. 2008;132:344–362. - PubMed
-
- Delhase M., Hayakawa M., Karin M. Positive and negative regulation of IκB kinase activity through IKKβ subunit phosphorylation. Science. 1999;284:309–313. - PubMed
-
- Palkowitsch L., Leidner J., Marienfeld R.B. Phosphorylation of serine 68 in the IκB kinase (IKK)-binding domain of NEMO interferes with the structure of the IKK complex and Tumor Necrosis Factor-α -induced NFκB activity. J. Biol. Chem. 2008;283:76–86. - PubMed
-
- Kray A.E., Carter R.S., Wadzinski B.E. Positive regulation of IκB kinase signaling by protein serine/threonine phosphatase 2A. J. Biol. Chem. 2005;280:35974–35982. - PubMed
-
- Broemer M., Krappmann D., Scheidereit C. Requirement of Hsp90 activity for IκB kinase (IKK) biosynthesis and for constitutive and inducible IKK and NF-κB activation. Oncogene. 2004;23:5378–5386. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
