

Thermodynamic origins of protein folding, allostery, and capsid formation in the human hepatitis B virus core protein
- PMID: 23824290
- PMCID: PMC3725087
- DOI: 10.1073/pnas.1308846110
Thermodynamic origins of protein folding, allostery, and capsid formation in the human hepatitis B virus core protein
Abstract
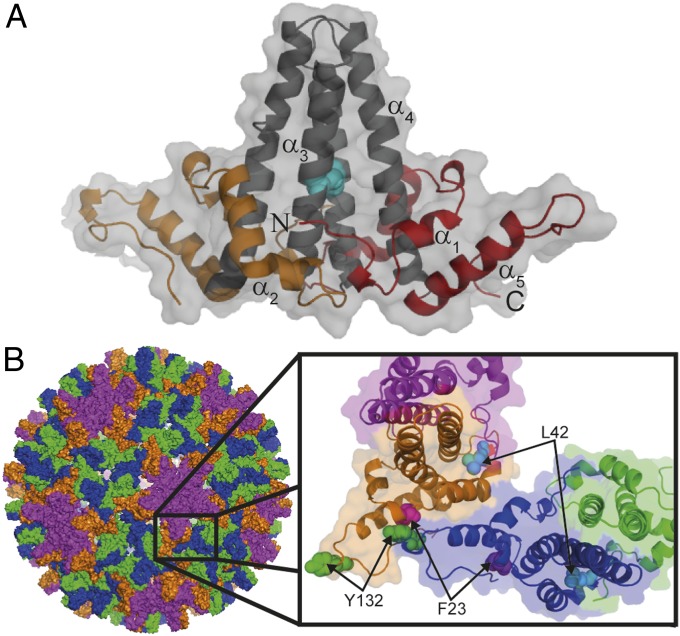
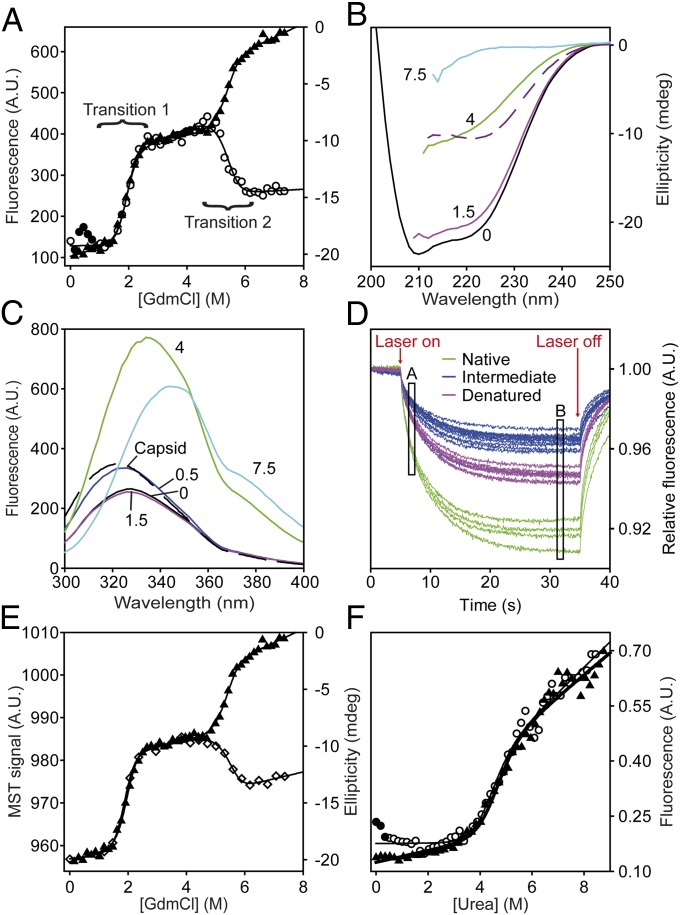
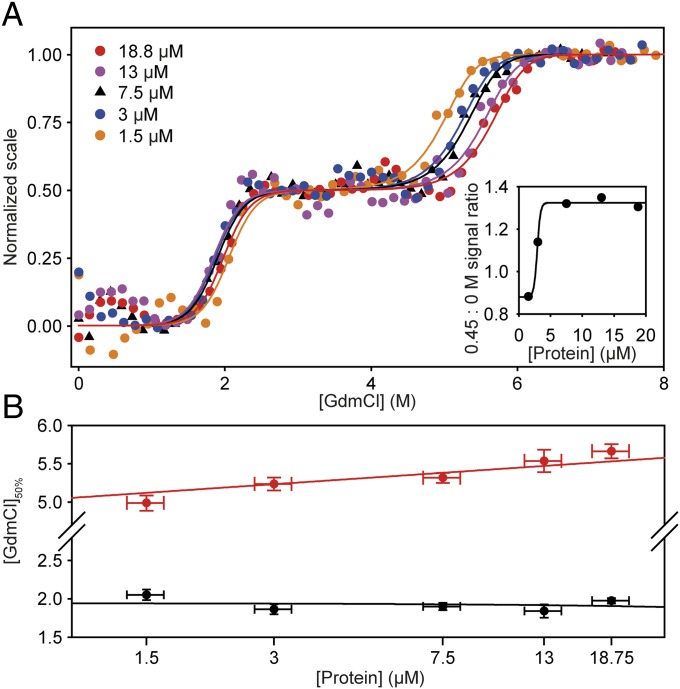
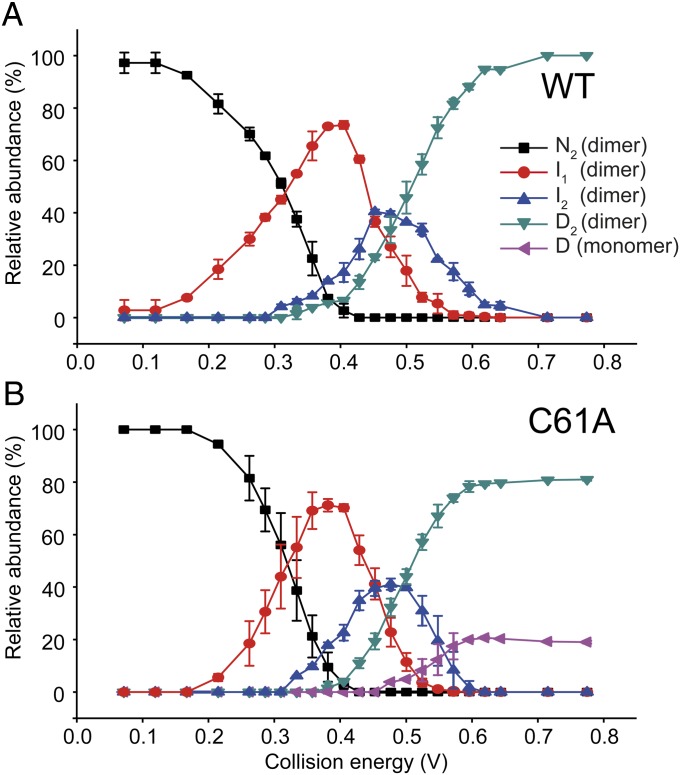
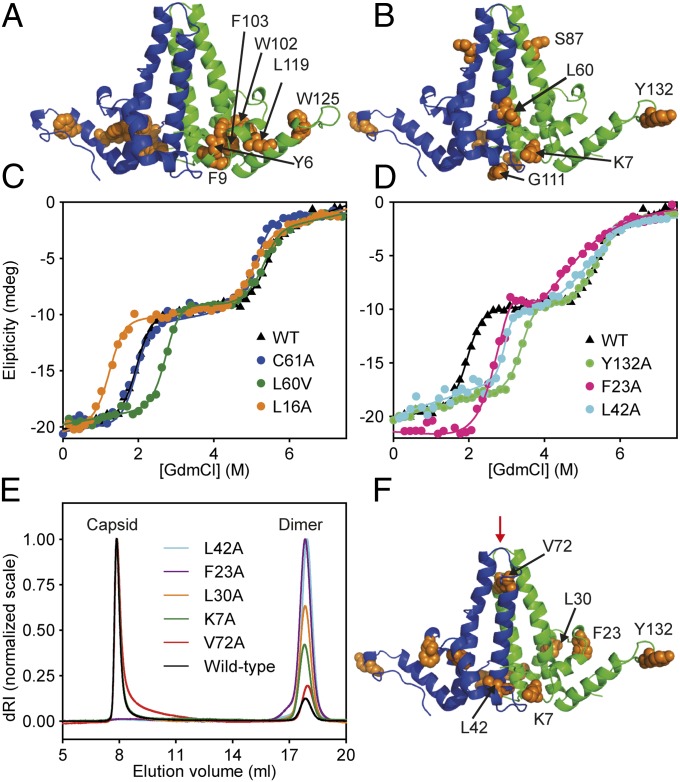
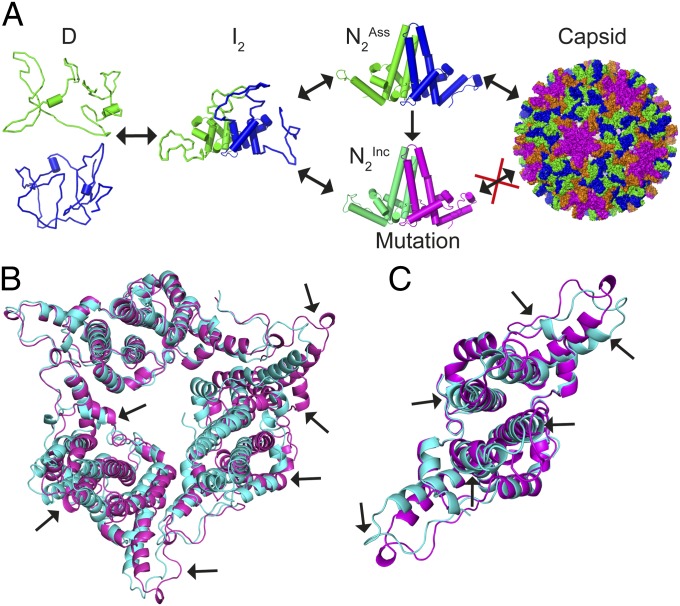
HBc, the capsid-forming "core protein" of human hepatitis B virus (HBV), is a multidomain, α-helical homodimer that aggressively forms human HBV capsids. Structural plasticity has been proposed to be important to the myriad functions HBc mediates during viral replication. Here, we report detailed thermodynamic analyses of the folding of the dimeric HBc protomer under conditions that prevented capsid formation. Central to our success was the use of ion mobility spectrometry-mass spectrometry and microscale thermophoresis, which allowed folding mechanisms to be characterized using just micrograms of protein. HBc folds in a three-state transition with a stable, dimeric, α-helical intermediate. Extensive protein engineering showed thermodynamic linkage between different structural domains. Unusual effects associated with mutating some residues suggest structural strain, arising from frustrated contacts, is present in the native dimer. We found evidence of structural gatekeepers that, when mutated, alleviated native strain and prevented (or significantly attenuated) capsid formation by tuning the population of alternative native conformations. This strain is likely an evolved feature that helps HBc access the different structures associated with its diverse essential functions. The subtle balance between native and strained contacts may provide the means to tune conformational properties of HBc by molecular interactions or mutations, thereby conferring allosteric regulation of structure and function. The ability to trap HBc conformers thermodynamically by mutation, and thereby ablate HBV capsid formation, provides proof of principle for designing antivirals that elicit similar effects.
Keywords: capsid assembly; energy landscape; protein dynamics; thermodynamic coupling.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Onuchic JN, Wolynes PG. Theory of protein folding. Curr Opin Struct Biol. 2004;14(1):70–75. - PubMed
-
- Jackson SE. How do small single-domain proteins fold? Fold Des. 1998;3(4):R81–R91. - PubMed
-
- Teague SJ. Implications of protein flexibility for drug discovery. Nat Rev Drug Discov. 2003;2(7):527–541. - PubMed
-
- World Health Organization Publication Hepatitis B vaccines: WHO position paper—Recommendations. Vaccine. 2010;28(3):589–590. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
