

Comparative Study
doi: 10.1128/JVI.01562-13.
Epub 2013 Jul 3.
Bilateral entry and release of Middle East respiratory syndrome coronavirus induces profound apoptosis of human bronchial epithelial cells
Affiliations
- PMID: 23824802
- PMCID: PMC3754134
- DOI: 10.1128/JVI.01562-13
Item in Clipboard
Comparative Study
Bilateral entry and release of Middle East respiratory syndrome coronavirus induces profound apoptosis of human bronchial epithelial cells
J Virol.
2013 Sep.
Abstract
The newly emerged Middle East respiratory syndrome coronavirus (MERS-CoV) infects human bronchial epithelial Calu-3 cells. Unlike severe acute respiratory syndrome (SARS)-CoV, which exclusively infects and releases through the apical route, this virus can do so through either side of polarized Calu-3 cells. Infection results in profound apoptosis within 24 h irrespective of its production of titers that are lower than those of SARS-CoV. Together, our results provide new insights into the dissemination and pathogenesis of MERS-CoV and may indicate that the virus differs markedly from SARS-CoV.
Figures
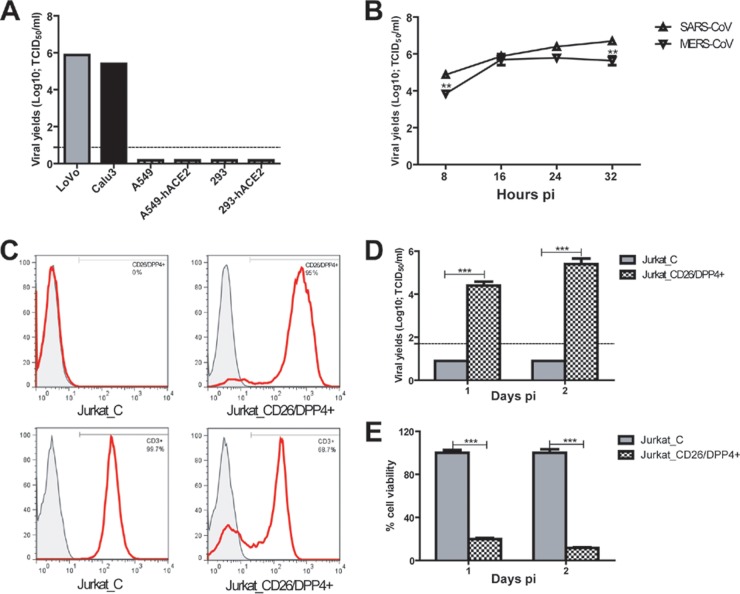
Human bronchial epithelial Calu-3 cells, colonic LoVo epithelial cells, and CD26/DPP4-expressing human Jurkat T cells are permissive for MERS-CoV/EMC-2012 infection. (A) Confluent Calu-3 cells, LoVo cells, alveolar A549 cells, ACE2-expressing A549 cells, embryonic kidney 293 cells, and ACE2-expressing 293 cells, grown in 12-well plates, were infected with MERS-CoV/EMC-2012 at an MOI of 0.1 for 24 h before harvesting of cell-free supernatants for assessing infectious viral titers by the standard Vero E6-based 50% tissue culture infective dose (TCID50) assay. (B) The “one-step” growth curves of MERS-CoV/EMC-2012 and SARS-CoV (Urbani) were established by infecting confluent Calu-3 cells with an MOI of 3, as described above. Cell-free supernatants were harvested at the indicated time points for determining the yields of infectious progeny viruses. (C) Phenotypes of Jurkat T cells transfected with control-versus-CD26/DPP4-encoding plasmid. (D) CD26/DPP4 expression confers the susceptibity of Jurkat T cells to productive MERS-CoV/EMC-2012 infection. (E) Trypan blue exclusion assay was used to simultaneously monitor the viability and mortality of infected Jurkat T cells. **, P < 0.01; ***, P < 0.001 (Student's t test). Data are representative of two independently conducted experiments.
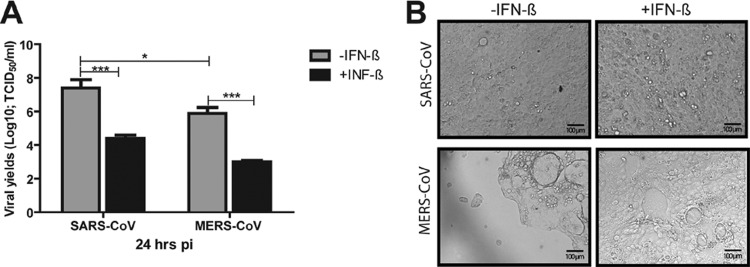
Prior IFN-β treatment effectively inhibits the replication of both SARS-CoV and MERS-CoV and attenuates MERS-CoV-induced CPE. Confluent monolayers of Calu-3 cells, grown in 12-well plates, were treated with recombinant (r) IFN-β for 16 h before being infected with either SARS-CoV or MERS-CoV/EMC-2012 (MOI = 0.1). The resulting cell-free supernatants were harvested at 24 h postinfection (pi) to determine the titers of infectious viruses as described for Fig. 1 (A), whereas the infected monolayer was fixed with paraformaldehyde (4%)–phosphate-buffered saline (PBS) for documentation of MERS-CoV-induced CPE (B). *, P < 0.05; ***, P < 0.001 (Student's t test).
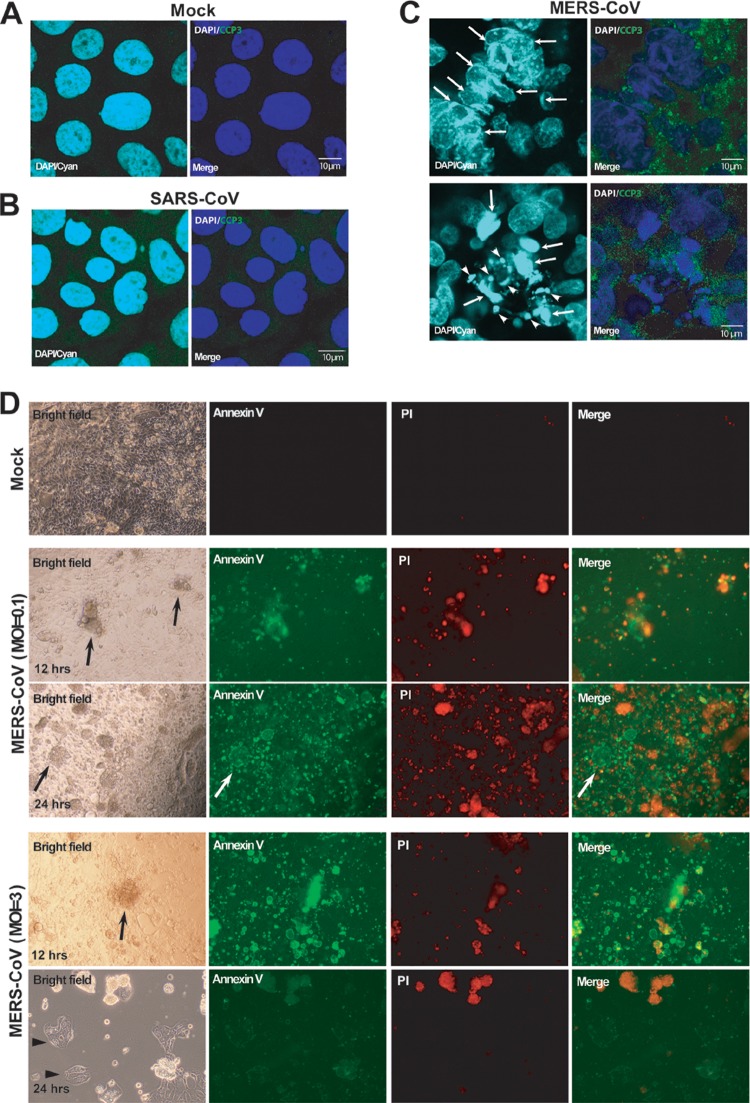
MERS-CoV, but not SARS-CoV, rapidly induces apoptosis of infected Calu-3 cells. Calu-3 cells grown in chamber slides were subjected to either SARS-CoV or MERS-CoV/EMC-2012 (MOI = 0.1) or remained uninfected for 24 h before we analyzed characteristic markers of apoptosis by confocal microscopy. Specifically, the morphology of the nuclei was analyzed after staining with 4′,6-diamidino-2-phenylindole (DAPI), whereas the expression of cleaved caspase 3 (CCP3) was evaluated after staining with Alexa 488-conjugated anti-CCP3 antibodies. The resulting specimens were examined using confocal microscopy at a high magnification. (A and B) Nuclei of mock-infected cells (A) or SARS-CoV-infected cells (B) possess a homogeneously round and smooth morphology without detectable CCP3 expression. (C) In contrast, we readily detected in MERS-CoV/EMC-2012-infected cells prominent nuclear margination, chromatin condensation (arrows), and typical apoptotic bodies (arrowheads) accompanied by positive staining for CCP3 (bright green) in the cytoplasm. (D) Additionally, FITC-conjugated annexin V and propidium iodide (PI) staining was used to discriminate between early apoptotic and late apoptotic/necrotic cells in MERS-CoV/EMC-2012-infected (MOI = 0.1 and 3) and uninfected Calu-3 cultures at both 12 and 24 h pi. Magnification, ×100.
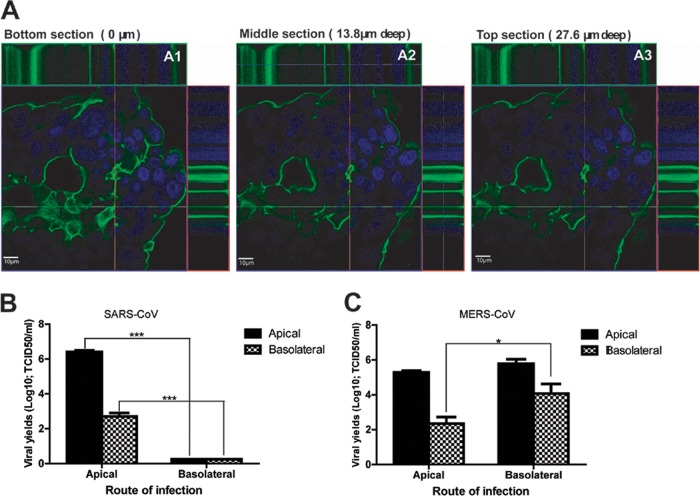
Unbiased apical and basoalteral expression of CD26/DPP4 enables bilateral MERS-CoV entry and release from polarized Calu-3 cells. (A) Confluent Calu-3 cells grown in the chamber slides were subjected to staining for CD26/DPP4 expression. The images of CD26/DPP4 surface expression (green) and nuclear staining (DAPI; blue) were captured by z-stack scanning from the bottom (A1), the middle (A2), and the top (A3) by using an inverted confocal microscope (Zeiss LSM 710). It is clear that CD26/DPP4 is uniformly expressed on the cell membrane of Calu-3 cells. (B and C) Highly polarized Calu-3 cells grown on filter inserts were inoculated with SARS-CoV (B) or MERS-CoV/EMC-2012 (C), at an MOI of 0.1, through either the apical or basolateral surface. At 24 h after infection, the culture supernatants collected from the apical and basolateral chambers were assessed for the contents of infectious viral particles by the TCID50 assay. *, P < 0.05; ***, P < 0.001 (Student's t test). The results are derived from representative findings for two independently conducted experiments.
References
-
- de Groot RJ, Baker SC, Baric RS, Brown CS, Drosten C, Enjuanes L, Fouchier RA, Galiano M, Gorbalenya AE, Memish Z, Perlman S, Poon LL, Snijder EJ, Stephens GM, Woo PC, Zaki AM, Zambon M, Ziebuhr J. 2013. Middle East respiratory syndrome coronavirus (MERS-CoV); announcement of the Coronavirus Study Group. J. Virol. 87:7790–7792 - PMC - PubMed
-
- WHO 22 May 2013. Novel coronavirus infection—update. WHO, Geneva, Switzerland: http://www.who.int/csr/don/2013_05_22_ncov/en/index.html
-
- van Boheemen S, de Graaf M, Lauber C, Bestebroer TM, Raj VS, Zaki AM, Osterhaus AD, Haagmans BL, Gorbalenya AE, Snijder EJ, Fouchier RA. 2012. Genomic characterization of a newly discovered coronavirus associated with acute respiratory distress syndrome in humans. mBio 3:e00473–12.10.1128/mBio.00473-12 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
