

Life as we know it
- PMID: 23825119
- PMCID: PMC3730701
- DOI: 10.1098/rsif.2013.0475
Life as we know it
Abstract
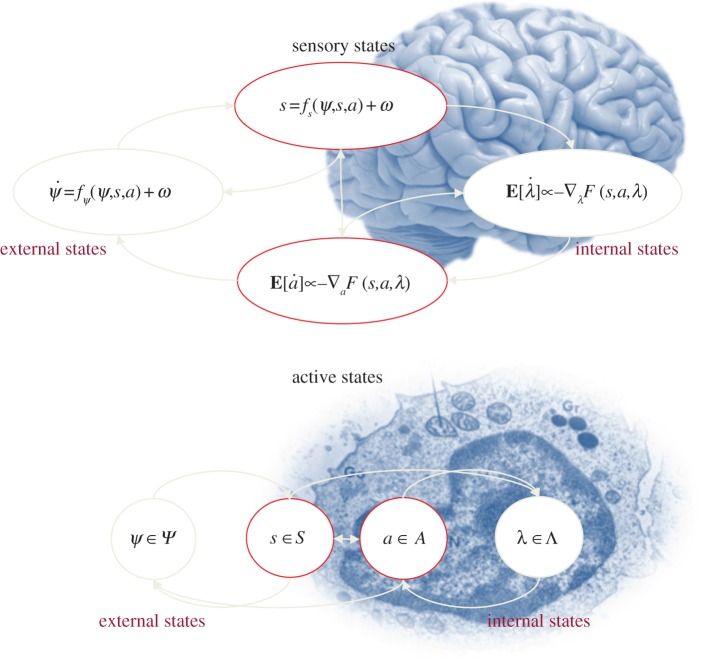
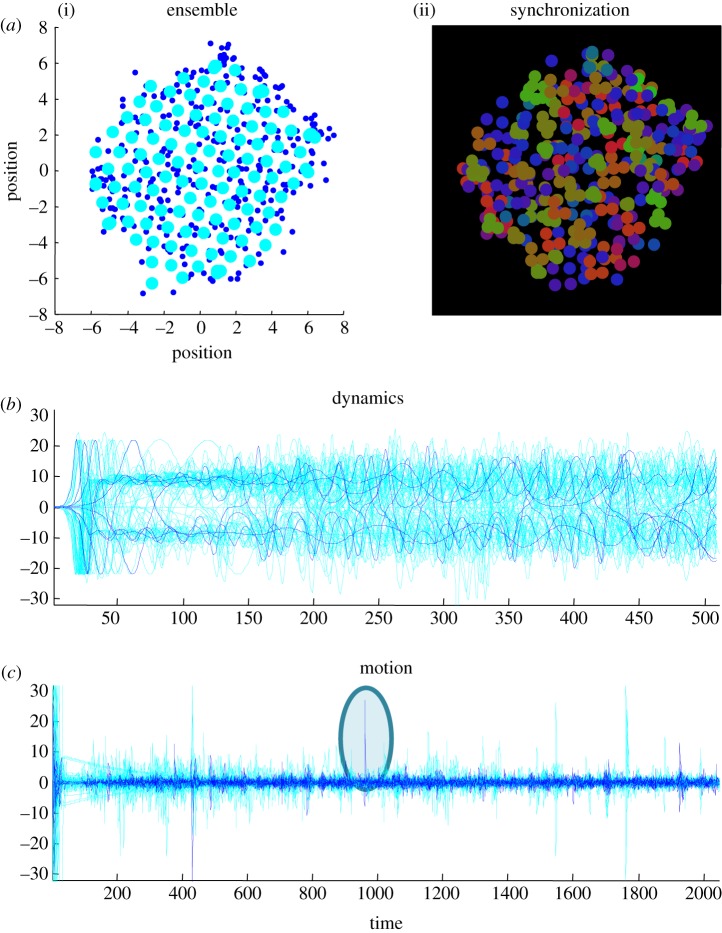
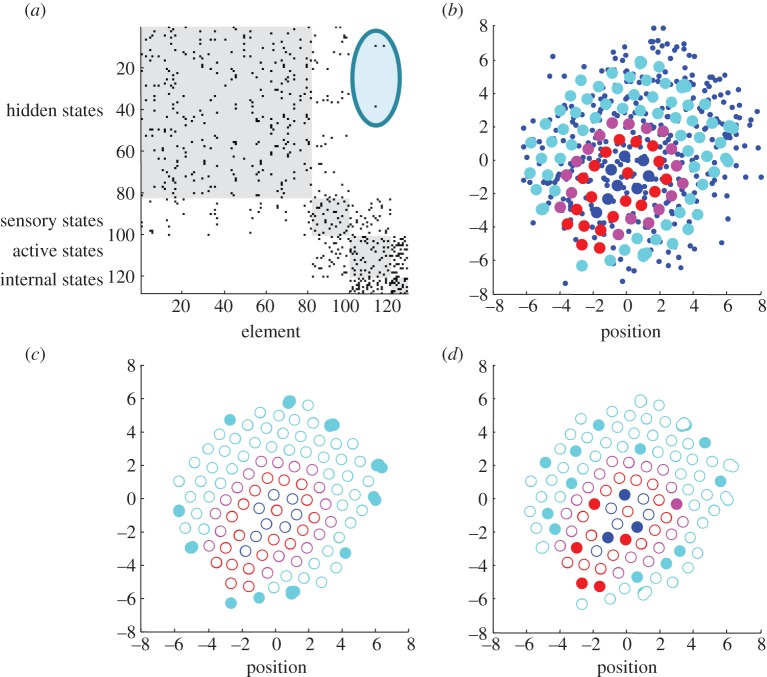
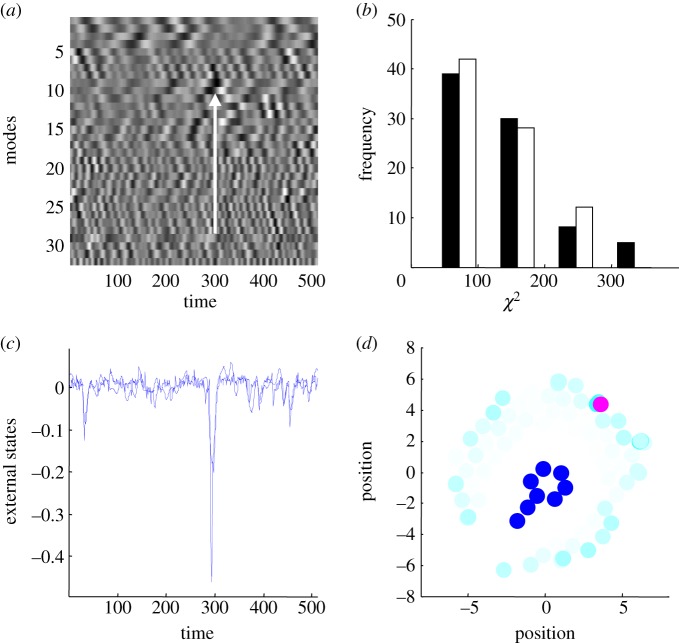
This paper presents a heuristic proof (and simulations of a primordial soup) suggesting that life-or biological self-organization-is an inevitable and emergent property of any (ergodic) random dynamical system that possesses a Markov blanket. This conclusion is based on the following arguments: if the coupling among an ensemble of dynamical systems is mediated by short-range forces, then the states of remote systems must be conditionally independent. These independencies induce a Markov blanket that separates internal and external states in a statistical sense. The existence of a Markov blanket means that internal states will appear to minimize a free energy functional of the states of their Markov blanket. Crucially, this is the same quantity that is optimized in Bayesian inference. Therefore, the internal states (and their blanket) will appear to engage in active Bayesian inference. In other words, they will appear to model-and act on-their world to preserve their functional and structural integrity, leading to homoeostasis and a simple form of autopoiesis.
Keywords: active inference; autopoiesis; ergodicity; free energy; random attractor; self-organization.
Figures
References
-
- Schrödinger E. 1944. What is life?: the physical aspect of the living cell. Dublin, Ireland: Trinity College.
-
- Haken H. 1983. Synergetics: an introduction. Non-equilibrium phase transition and self-selforganisation in physics, chemistry and biology, 3rd edn Berlin, Germany: Springer.
-
- Maturana HR, Varela F. (eds) 1980. Autopoiesis and cognition. Dordrecht, The Netherlands: Reidel.
-
- Nicolis G, Prigogine I. 1977. Self-organization in non-equilibrium systems. New York, NY: Wiley.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
