

Animal activity around the clock with no overt circadian rhythms: patterns, mechanisms and adaptive value
- PMID: 23825202
- PMCID: PMC3712434
- DOI: 10.1098/rspb.2013.0019
Animal activity around the clock with no overt circadian rhythms: patterns, mechanisms and adaptive value
Abstract
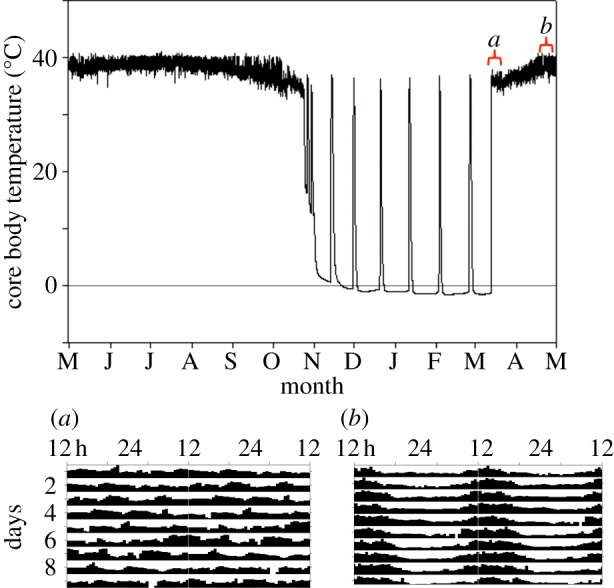
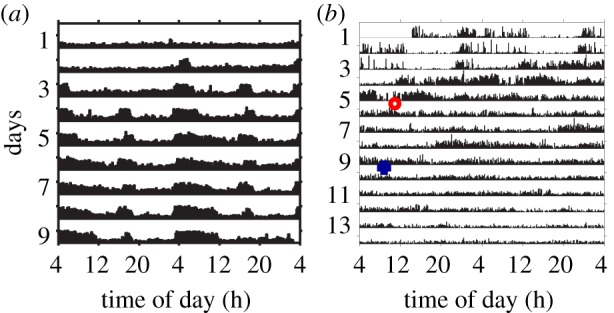
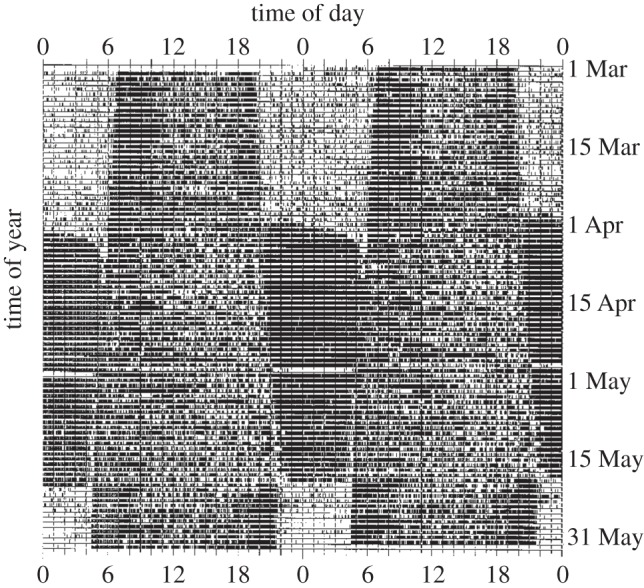
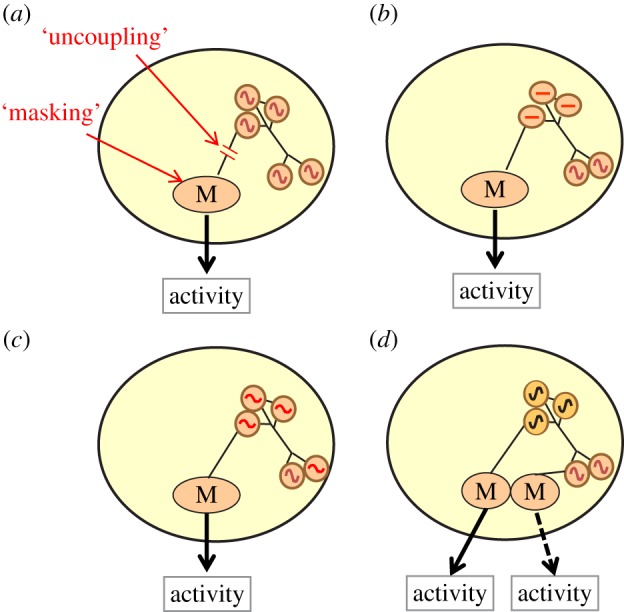
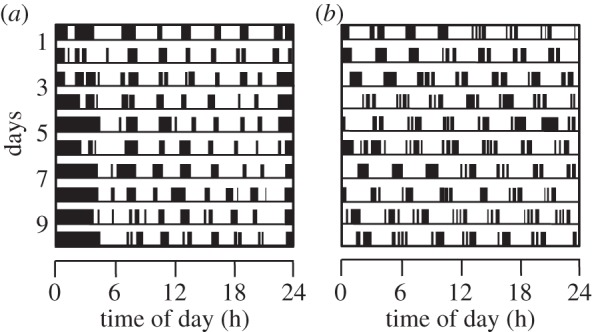
Circadian rhythms are ubiquitous in many organisms. Animals that are forced to be active around the clock typically show reduced performance, health and survival. Nevertheless, we review evidence of animals showing prolonged intervals of activity with attenuated or nil overt circadian rhythms and no apparent ill effects. We show that around-the-clock and ultradian activity patterns are more common than is generally appreciated, particularly in herbivores, in animals inhabiting polar regions and habitats with constant physical environments, in animals during specific life-history stages (such as migration or reproduction), and in highly social animals. The underlying mechanisms are diverse, but studies suggest that some circadian pacemakers continue to measure time in animals active around the clock. The prevalence of around-the-clock activity in diverse animals and habitats, and an apparent diversity of underlying mechanisms, are consistent with convergent evolution. We suggest that the basic organizational principles of the circadian system and its complexity encompass the potential for chronobiological plasticity. There may be trade-offs between benefits of persistent daily rhythms versus plasticity, which for reasons still poorly understood make overt daily arrhythmicity functionally adaptive only in selected habitats and for selected lifestyles.
Keywords: arrhythmic; circadian; evolution; plasticity; sleep; ultradian rhythms.
Figures
References
-
- Foster RG, Kreitzman L. 2005. Rhythms of life: the biological clocks that control the daily lives of every living thing. New Haven, CT: Yale University Press
-
- Herzog ED. 2007. Neurons and networks in daily rhythms. Nat. Rev. Neurosci. 8, 790–802 (doi:10.1038/nrn2215) - DOI - PubMed
-
- O'Neill JS, Reddy AB. 2011. Circadian clocks in human red blood cells. Nature 469, 498–503 (doi:10.1038/nature09702) - DOI - PMC - PubMed
-
- Menaker M. 2006. Circadian organization in the real world. Proc. Natl Acad. Sci. USA 103, 3015–3016 (doi:10.1073/pnas.0600360103) - DOI - PMC - PubMed
-
- Van der Zee EA, Havekes R, Barf RP, Hut RA, Nijholt IM, Jacobs EH, Gerkema MP. 2008. Circadian time-place learning in mice depends on Cry genes. Curr. Biol. 18, 844–848 (doi:10.1016/j.cub.2008.04.077) - DOI - PubMed
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
