

edn1 and hand2 Interact in early regulation of pharyngeal arch outgrowth during zebrafish development
- PMID: 23826316
- PMCID: PMC3691169
- DOI: 10.1371/journal.pone.0067522
edn1 and hand2 Interact in early regulation of pharyngeal arch outgrowth during zebrafish development
Abstract
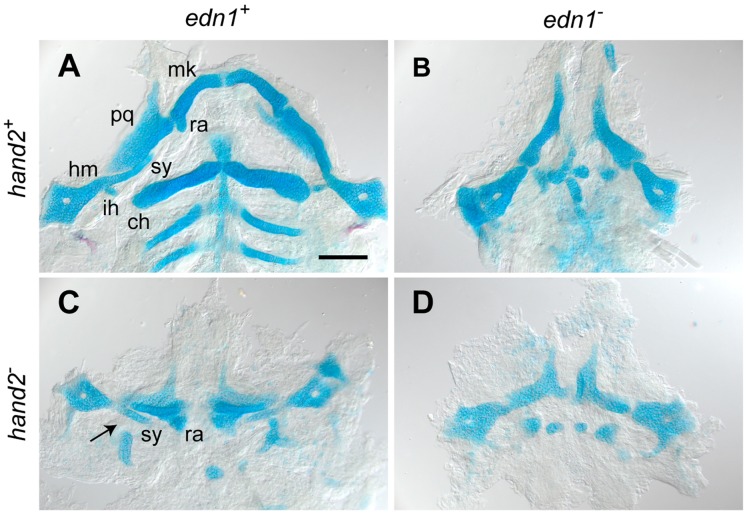
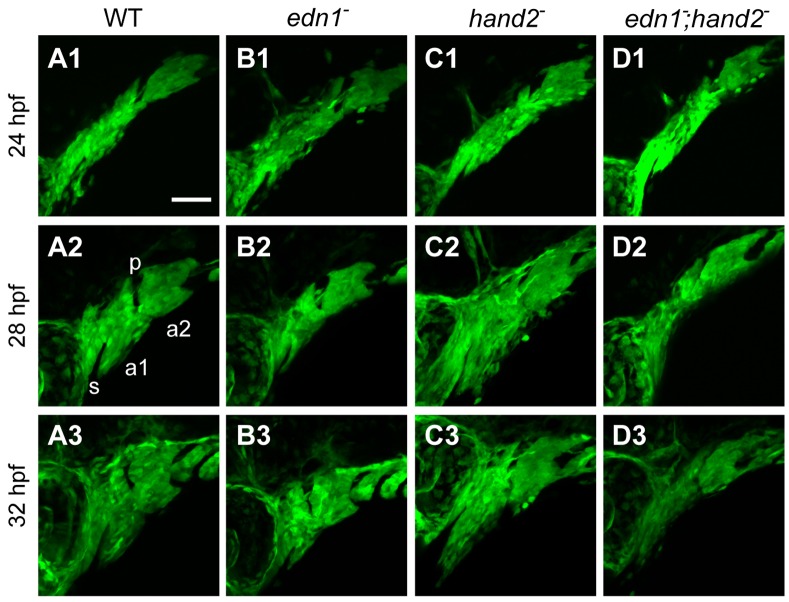
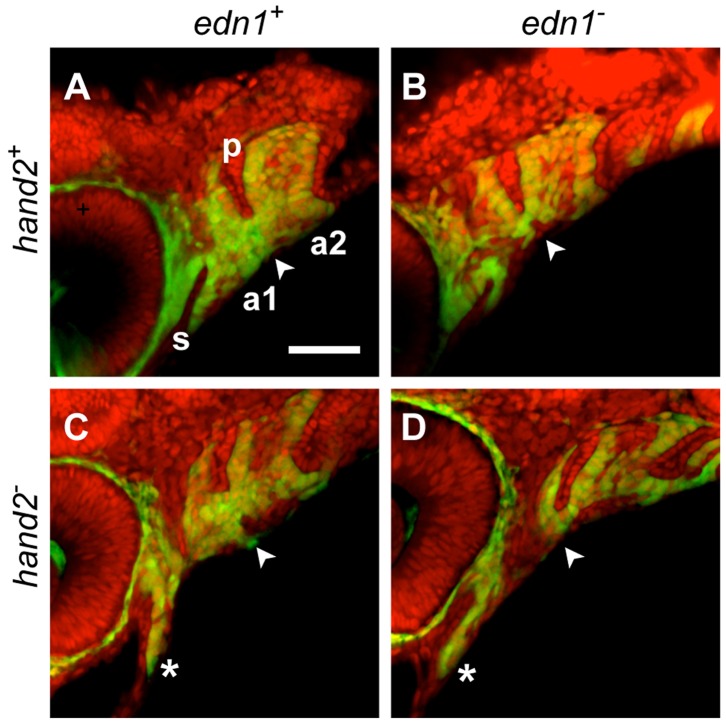
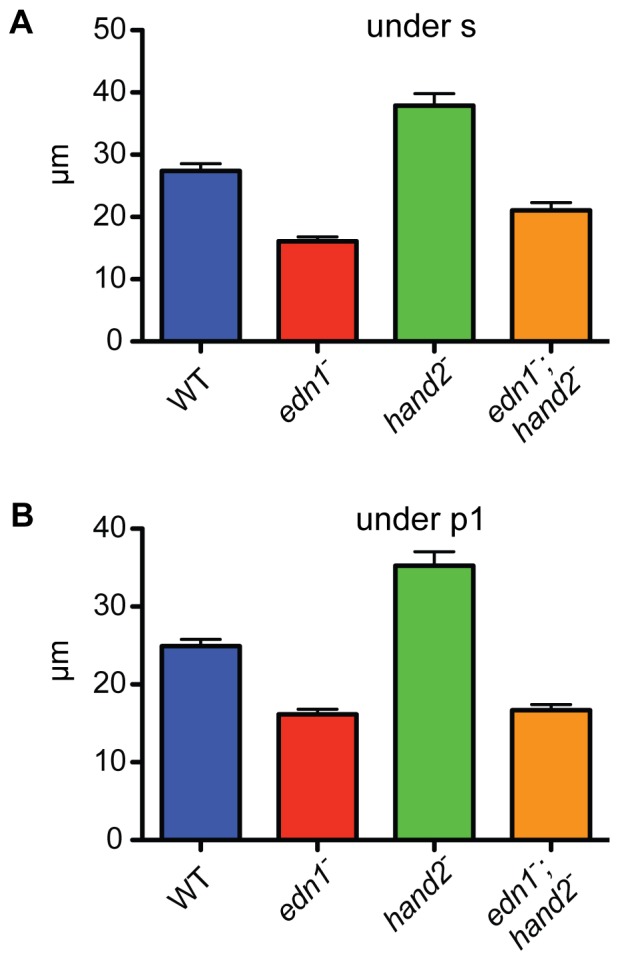
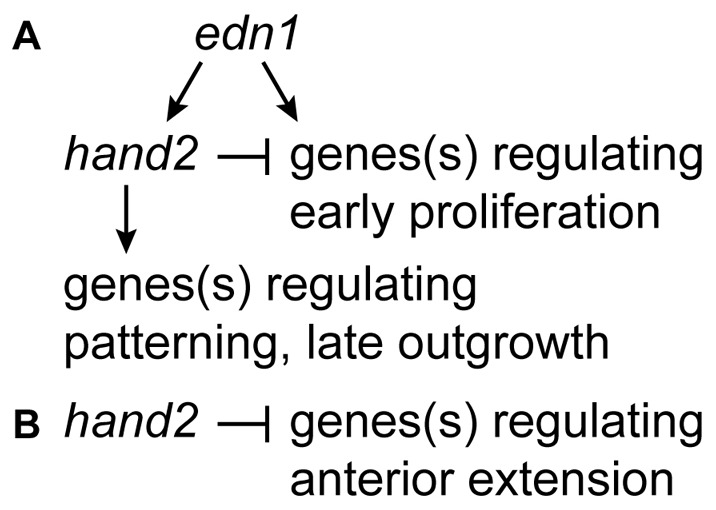
Endothelin-1 (Edn1) signaling provides a critical input to development of the embryonic pharygneal arches and their skeletal derivatives, particularly the articulating joints and the ventral skeleton including the lower jaw. Previous work in zebrafish has mostly focused on the role of Edn1 in dorsal-ventral (DV) patterning, but Edn1 signaling must also regulate tissue size, for with severe loss of the pathway the ventral skeleton is not only mispatterned, but is also prominently hypoplastic--reduced in size. Here we use mutational analyses to show that in the early pharyngeal arches, ventral-specific edn1-mediated proliferation of neural crest derived cells is required for DV expansion and outgrowth, and that this positive regulation is counterbalanced by a negative one exerted through a pivotal, ventrally expressed Edn1-target gene, hand2. We also describe a new morphogenetic cell movement in the ventral first arch, sweeping cells anterior in the arch to the region where the lower jaw forms. This movement is negatively regulated by hand2 in an apparently edn1-independent fashion. These findings point to complexity of regulation by edn1 and hand2 at the earliest stages of pharyngeal arch development, in which control of growth and morphogenesis can be genetically separated.
Conflict of interest statement
Figures
Similar articles
-
Two endothelin 1 effectors, hand2 and bapx1, pattern ventral pharyngeal cartilage and the jaw joint.Development. 2003 Apr;130(7):1353-65. doi: 10.1242/dev.00339. Development. 2003. PMID: 12588851
-
Combinatorial roles for BMPs and Endothelin 1 in patterning the dorsal-ventral axis of the craniofacial skeleton.Development. 2011 Dec;138(23):5135-46. doi: 10.1242/dev.067801. Epub 2011 Oct 26. Development. 2011. PMID: 22031543 Free PMC article.
-
Wnt signaling interacts with bmp and edn1 to regulate dorsal-ventral patterning and growth of the craniofacial skeleton.PLoS Genet. 2014 Jul 24;10(7):e1004479. doi: 10.1371/journal.pgen.1004479. eCollection 2014 Jul. PLoS Genet. 2014. PMID: 25058015 Free PMC article.
-
Regulation of facial morphogenesis by endothelin signaling: insights from mice and fish.Am J Med Genet A. 2010 Dec;152A(12):2962-73. doi: 10.1002/ajmg.a.33568. Am J Med Genet A. 2010. PMID: 20684004 Free PMC article. Review.
-
Understanding endothelin-1 function during craniofacial development in the mouse and zebrafish.Birth Defects Res C Embryo Today. 2004 Jun;72(2):190-9. doi: 10.1002/bdrc.20007. Birth Defects Res C Embryo Today. 2004. PMID: 15269892 Review.
Cited by
-
Dlx5 and Dlx6 can antagonize cell division at the G1/S checkpoint.BMC Mol Cell Biol. 2019 Apr 11;20(1):8. doi: 10.1186/s12860-019-0191-6. BMC Mol Cell Biol. 2019. PMID: 31041891 Free PMC article.
-
The Wnt Co-Receptor Lrp5 Is Required for Cranial Neural Crest Cell Migration in Zebrafish.PLoS One. 2015 Jun 29;10(6):e0131768. doi: 10.1371/journal.pone.0131768. eCollection 2015. PLoS One. 2015. PMID: 26121341 Free PMC article.
-
Transgene-mediated skeletal phenotypic variation in zebrafish.J Fish Biol. 2021 Apr;98(4):956-970. doi: 10.1111/jfb.14300. Epub 2020 Mar 30. J Fish Biol. 2021. PMID: 32112658 Free PMC article.
-
Terminal chromosome 4q deletion syndrome in an infant with hearing impairment and moderate syndromic features: review of literature.BMC Med Genet. 2014 Jun 25;15:72. doi: 10.1186/1471-2350-15-72. BMC Med Genet. 2014. PMID: 24962056 Free PMC article. Review.
-
Variable paralog expression underlies phenotype variation.Elife. 2022 Sep 22;11:e79247. doi: 10.7554/eLife.79247. Elife. 2022. PMID: 36134886 Free PMC article.
References
-
- Schilling TF, Piotrowski T, Grandel H, Brand M, Heisenberg CP, et al. (1996) Jaw and branchial arch mutants in zebrafish I: branchial arches. Development 123: 329–344. - PubMed
-
- Piotrowski T, Schilling TF, Brand M, Jiang YJ, Heisenberg CP, et al. (1996) Jaw and branchial arch mutants in zebrafish II: anterior arches and cartilage differentiation. Development 123: 345–356. - PubMed
-
- Clouthier DE, Schilling TF (2004) Understanding endothelin-1 function during craniofacial development in the mouse and zebrafish. Birth Defects Res C Embryo Today 72: 190–199. - PubMed
-
- Kimmel CB, Walker MB, Miller CT (2007) Morphing the hyomandibular skeleton in development and evolution. J Exp Zool B Mol Dev Evol 308: 609–624. - PubMed
-
- Thomas T, Kurihara H, Yamagishi H, Kurihara Y, Yazaki Y, et al. (1998) A signaling cascade involving endothelin-1, dHAND and Msx1 regulates development of neural-crest-derived branchial arch mesenchyme. Development 125: 3005–3014. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
