

Second cistron in CACNA1A gene encodes a transcription factor mediating cerebellar development and SCA6
- PMID: 23827678
- PMCID: PMC3939801
- DOI: 10.1016/j.cell.2013.05.059
Second cistron in CACNA1A gene encodes a transcription factor mediating cerebellar development and SCA6
Abstract
The CACNA1A gene, encoding the voltage-gated calcium channel subunit α1A, is involved in pre- and postsynaptic Ca(2+) signaling, gene expression, and several genetic neurological disorders. We found that CACNA1A coordinates gene expression using a bicistronic mRNA bearing a cryptic internal ribosomal entry site (IRES). The first cistron encodes the well-characterized α1A subunit. The second expresses a transcription factor, α1ACT, which coordinates expression of a program of genes involved in neural and Purkinje cell development. α1ACT also contains the polyglutamine (polyQ) tract that, when expanded, causes spinocerebellar ataxia type 6 (SCA6). When expressed as an independent polypeptide, α1ACT-bearing an expanded polyQ tract-lacks transcription factor function and neurite outgrowth properties, causes cell death in culture, and leads to ataxia and cerebellar atrophy in transgenic mice. Suppression of CACNA1A IRES function in SCA6 may be a potential therapeutic strategy.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures

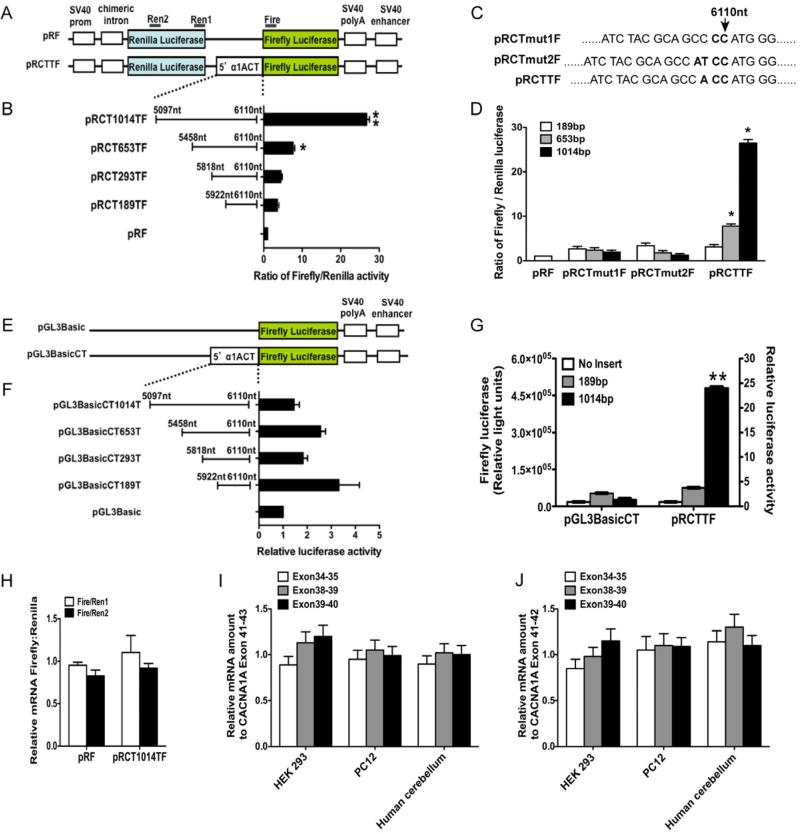



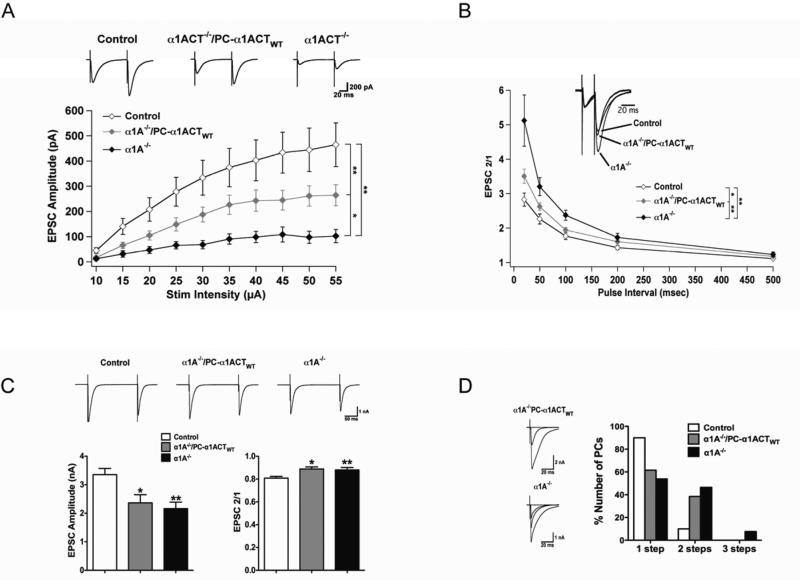
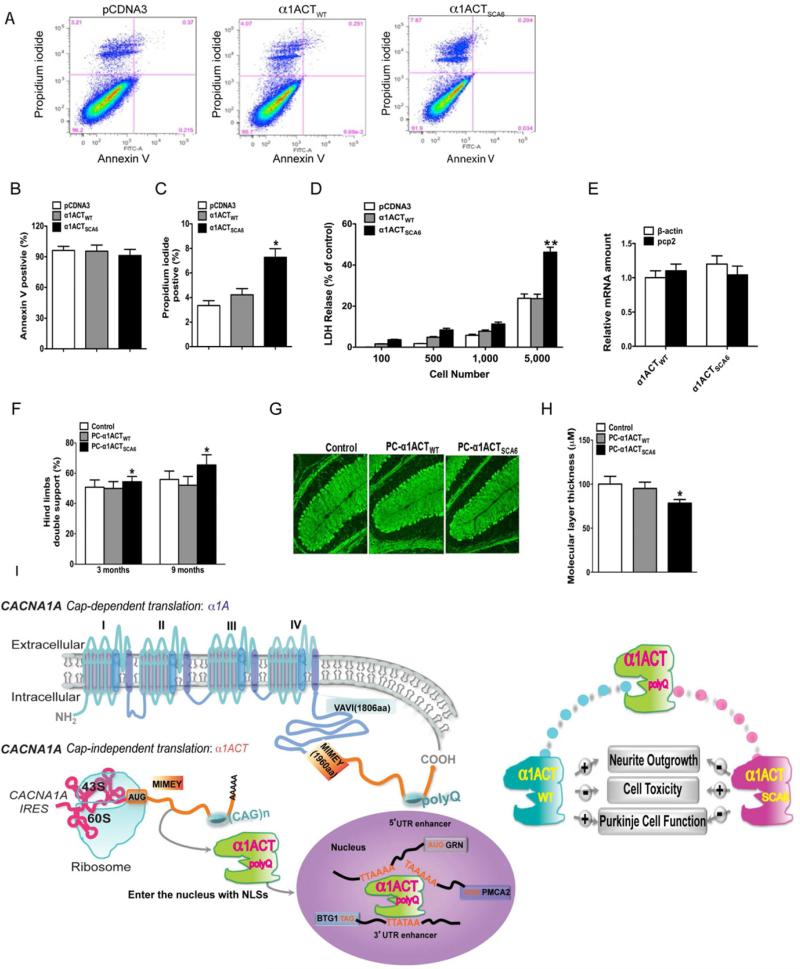
Comment in
-
Revelations from a bicistronic calcium channel gene.Cell Cycle. 2014;13(6):875-6. doi: 10.4161/cc.28199. Epub 2014 Feb 14. Cell Cycle. 2014. PMID: 24552819 Free PMC article. No abstract available.
References
-
- Baker M, Mackenzie IR, Pickering-Brown SM, Gass J, Rademakers R, Lindholm C, Snowden J, Adamson J, Sadovnick AD, Rollinson S, et al. Mutations in progranulin cause tau-negative frontotemporal dementia linked to chromosome 17. Nature. 2006;442:916–919. - PubMed
-
- Berthet C, Guehenneux F, Revol V, Samarut C, Lukaszewicz A, Dehay C, Dumontet C, Magaud JP, Rouault JP. Interaction of PRMT1 with BTG/TOB proteins in cell signalling: molecular analysis and functional aspects. Genes to cells : devoted to molecular & cellular mechanisms. 2002;7:29–39. - PubMed
-
- Cain SM, Snutch TP. Voltage-gated calcium channels and disease. Biofactors. 2011;37:197–205. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
