

A bacterial virulence protein promotes pathogenicity by inhibiting the bacterium's own F1Fo ATP synthase
- PMID: 23827679
- PMCID: PMC3736803
- DOI: 10.1016/j.cell.2013.06.004
A bacterial virulence protein promotes pathogenicity by inhibiting the bacterium's own F1Fo ATP synthase
Abstract
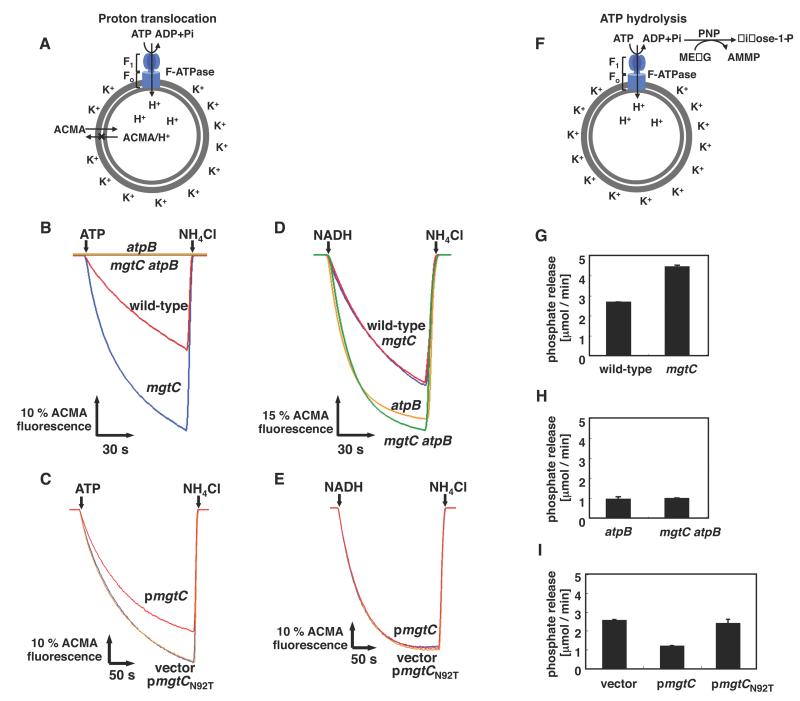
Several intracellular pathogens, including Salmonella enterica and Mycobacterium tuberculosis, require the virulence protein MgtC to survive within macrophages and to cause a lethal infection in mice. We now report that, unlike secreted virulence factors that target the host vacuolar ATPase to withstand phagosomal acidity, the MgtC protein acts on Salmonella's own F1Fo ATP synthase. This complex couples proton translocation to ATP synthesis/hydrolysis and is required for virulence. We establish that MgtC interacts with the a subunit of the F1Fo ATP synthase, hindering ATP-driven proton translocation and NADH-driven ATP synthesis in inverted vesicles. An mgtC null mutant displays heightened ATP levels and an acidic cytoplasm, whereas mgtC overexpression decreases ATP levels. A single amino acid substitution in MgtC that prevents binding to the F1Fo ATP synthase abolishes control of ATP levels and attenuates pathogenicity. MgtC provides a singular example of a virulence protein that promotes pathogenicity by interfering with another virulence protein.
Copyright © 2013 Elsevier Inc. All rights reserved.
Figures






Similar articles
-
The Salmonella virulence protein MgtC promotes phosphate uptake inside macrophages.Nat Commun. 2019 Jul 25;10(1):3326. doi: 10.1038/s41467-019-11318-2. Nat Commun. 2019. PMID: 31346161 Free PMC article.
-
Control of a Salmonella virulence locus by an ATP-sensing leader messenger RNA.Nature. 2012 Jun 13;486(7402):271-5. doi: 10.1038/nature11090. Nature. 2012. PMID: 22699622 Free PMC article.
-
Regulation and function of the Salmonella MgtC virulence protein.J Microbiol. 2015 Oct;53(10):667-72. doi: 10.1007/s12275-015-5283-1. Epub 2015 Aug 1. J Microbiol. 2015. PMID: 26231375 Review.
-
Peptide-assisted degradation of the Salmonella MgtC virulence factor.EMBO J. 2008 Feb 6;27(3):546-57. doi: 10.1038/sj.emboj.7601983. Epub 2008 Jan 17. EMBO J. 2008. PMID: 18200043 Free PMC article.
-
Unique structural and mechanistic properties of mycobacterial F-ATP synthases: Implications for drug design.Prog Biophys Mol Biol. 2020 May;152:64-73. doi: 10.1016/j.pbiomolbio.2019.11.006. Epub 2019 Nov 16. Prog Biophys Mol Biol. 2020. PMID: 31743686 Review.
Cited by
-
A protein that controls the onset of a Salmonella virulence program.EMBO J. 2018 Jul 13;37(14):e96977. doi: 10.15252/embj.201796977. Epub 2018 Jun 1. EMBO J. 2018. PMID: 29858228 Free PMC article.
-
Whole-Genome Comparative and Pathogenicity Analysis of Salmonella enterica subsp. enterica Serovar Rissen.G3 (Bethesda). 2020 Jul 7;10(7):2159-2170. doi: 10.1534/g3.120.401201. G3 (Bethesda). 2020. PMID: 32358017 Free PMC article.
-
Reduction in adaptor amounts establishes degradation hierarchy among protease substrates.Proc Natl Acad Sci U S A. 2018 May 8;115(19):E4483-E4492. doi: 10.1073/pnas.1722246115. Epub 2018 Apr 23. Proc Natl Acad Sci U S A. 2018. PMID: 29686082 Free PMC article.
-
Assessment of the requirements for magnesium transporters in Bacillus subtilis.J Bacteriol. 2014 Mar;196(6):1206-14. doi: 10.1128/JB.01238-13. Epub 2014 Jan 10. J Bacteriol. 2014. PMID: 24415722 Free PMC article.
-
Elongation factor P restricts Salmonella's growth by controlling translation of a Mg2+ transporter gene during infection.Sci Rep. 2017 Feb 9;7:42098. doi: 10.1038/srep42098. Sci Rep. 2017. PMID: 28181542 Free PMC article.
References
-
- Andries K, Verhasselt P, Guillemont J, Gohlmann HW, Neefs JM, Winkler H, Van Gestel J, Timmerman P, Zhu M, Lee E, et al. A diarylquinoline drug active on the ATP synthase of Mycobacterium tuberculosis. Science. 2005;307:223–227. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
