

Studies of efficacy and liver toxicity related to adeno-associated virus-mediated RNA interference
- PMID: 23829557
- PMCID: PMC3747597
- DOI: 10.1089/hum.2012.239
Studies of efficacy and liver toxicity related to adeno-associated virus-mediated RNA interference
Abstract
Adeno-associated virus (AAV)-mediated RNA interference shows promise as a therapy for chronic hepatitis B virus (HBV) infection, but its low efficacy and hepatotoxicity pose major challenges. We have generated AAV vectors containing different promoters and a panel of HBV-specific short hairpin RNAs (shRNAs) to investigate factors that contribute to the efficacy and pathogenesis of AAV-mediated RNA interference. HBV transgenic mice injected with high doses of AAV vectors containing the U6 promoter produced abundant shRNAs, transiently inhibited HBV, but induced severe hepatotoxicity. Sustained HBV suppression without liver toxicity can be achieved by lowering the dose of AAV-U6 vectors. AAVs containing the weaker H1 promoter did not cause liver injury, but their therapeutic efficacy was highly dependent on the sequence of the shRNA. Mice treated with the toxic U6-promoter-driven shRNA showed little change in hepatic microRNA levels, but a dramatic increase in hepatic leukocytes and inflammatory cytokines and chemokines. Hepatotoxicity was completely absent in immunodeficient mice and significantly alleviated in wild-type mice depleted of macrophages and granulocytes, suggesting that host inflammatory responses are the major cause of liver injury induced by the overexpressed shRNAs from AAV-U6 vectors. Our results demonstrate that selection of a highly potent shRNA and control its expression level is critical to achieve sustained HBV suppression without inducing inflammatory side effects.
Figures
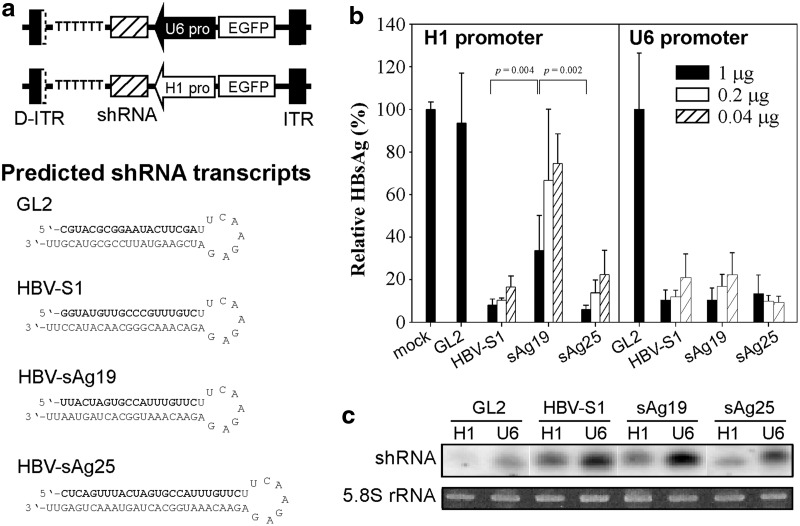
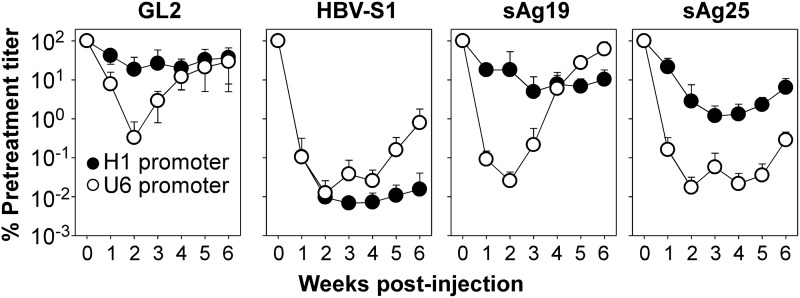
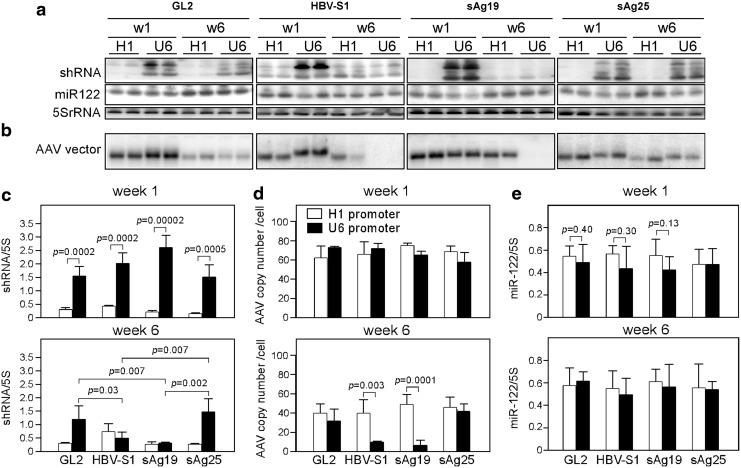
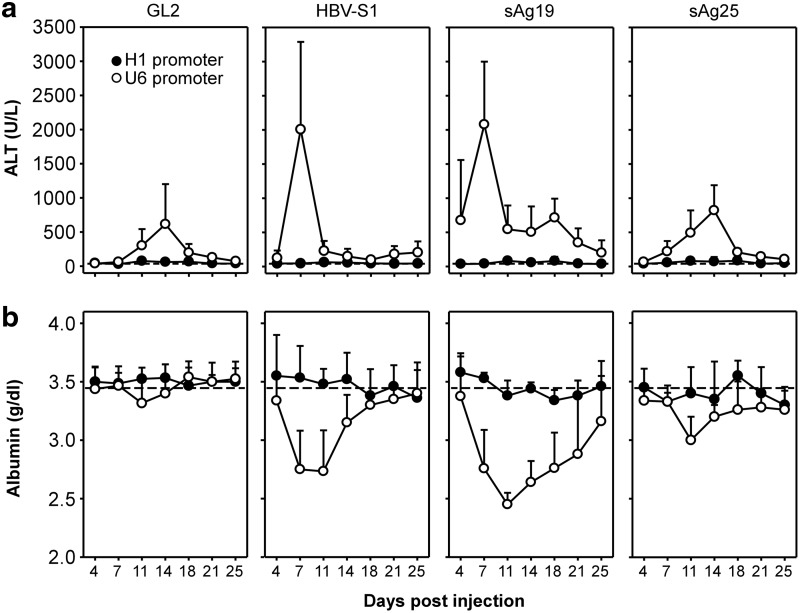
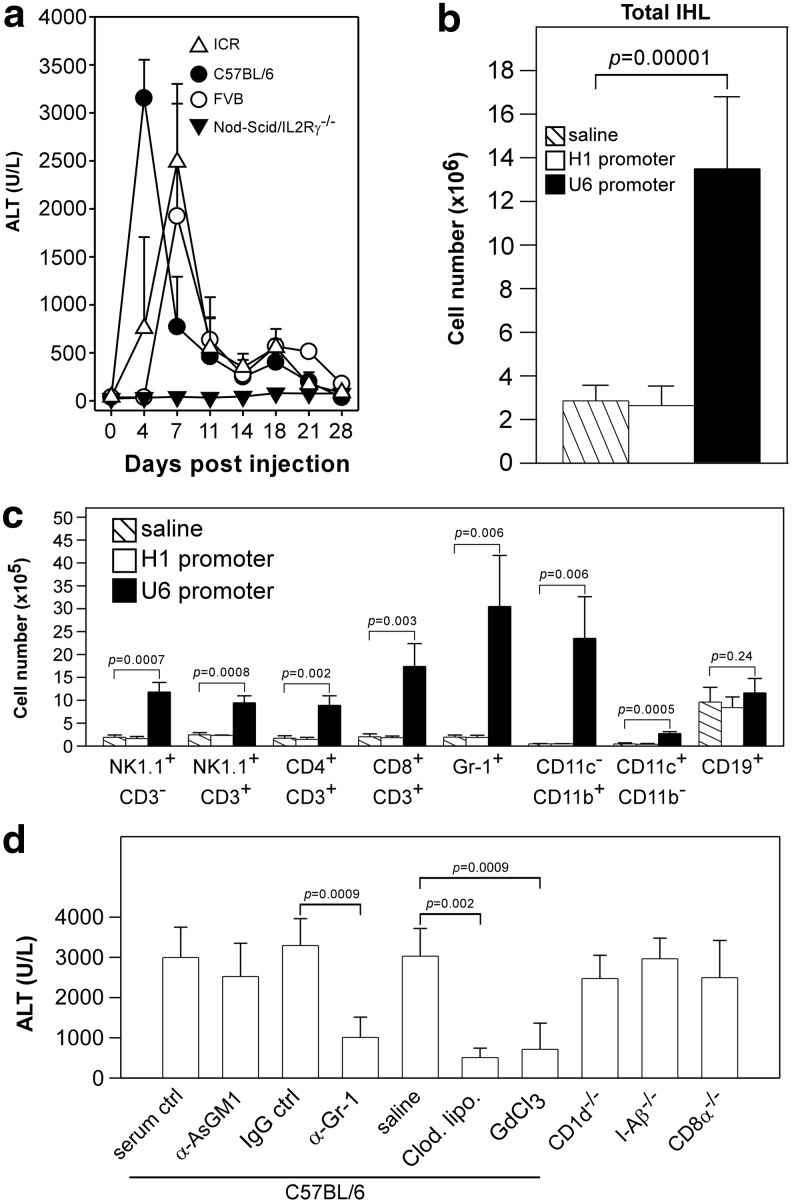
 , Saline; □, AAV8-H1/sAg19; ■, AAV8-U6/sAg19. The number of cells per liver for individual cell types was calculated by multiplying the percentage of the individual cell type by the total number of isolated liver leukocytes per liver. The experiment was performed twice with similar results. (d) Groups of wild-type C57BL/6 mice treated with anti-asialo-GM1 antiserum, anti-Gr-1 monoclonal antibody, clodronate liposome or GdCl3, CD1d−/− mice, I-Aβ−/− mice, or CD8−/− mice (n=5–8) were injected with AAV8-U6/sAg19 on day 0, and then serum samples were collected at day 4 for analysis of ALT activity (mean±SD). Wild-type C57BL/6 mice treated with control rabbit antiserum, control rat immunoglobulin G, or saline were included as controls.
, Saline; □, AAV8-H1/sAg19; ■, AAV8-U6/sAg19. The number of cells per liver for individual cell types was calculated by multiplying the percentage of the individual cell type by the total number of isolated liver leukocytes per liver. The experiment was performed twice with similar results. (d) Groups of wild-type C57BL/6 mice treated with anti-asialo-GM1 antiserum, anti-Gr-1 monoclonal antibody, clodronate liposome or GdCl3, CD1d−/− mice, I-Aβ−/− mice, or CD8−/− mice (n=5–8) were injected with AAV8-U6/sAg19 on day 0, and then serum samples were collected at day 4 for analysis of ALT activity (mean±SD). Wild-type C57BL/6 mice treated with control rabbit antiserum, control rat immunoglobulin G, or saline were included as controls.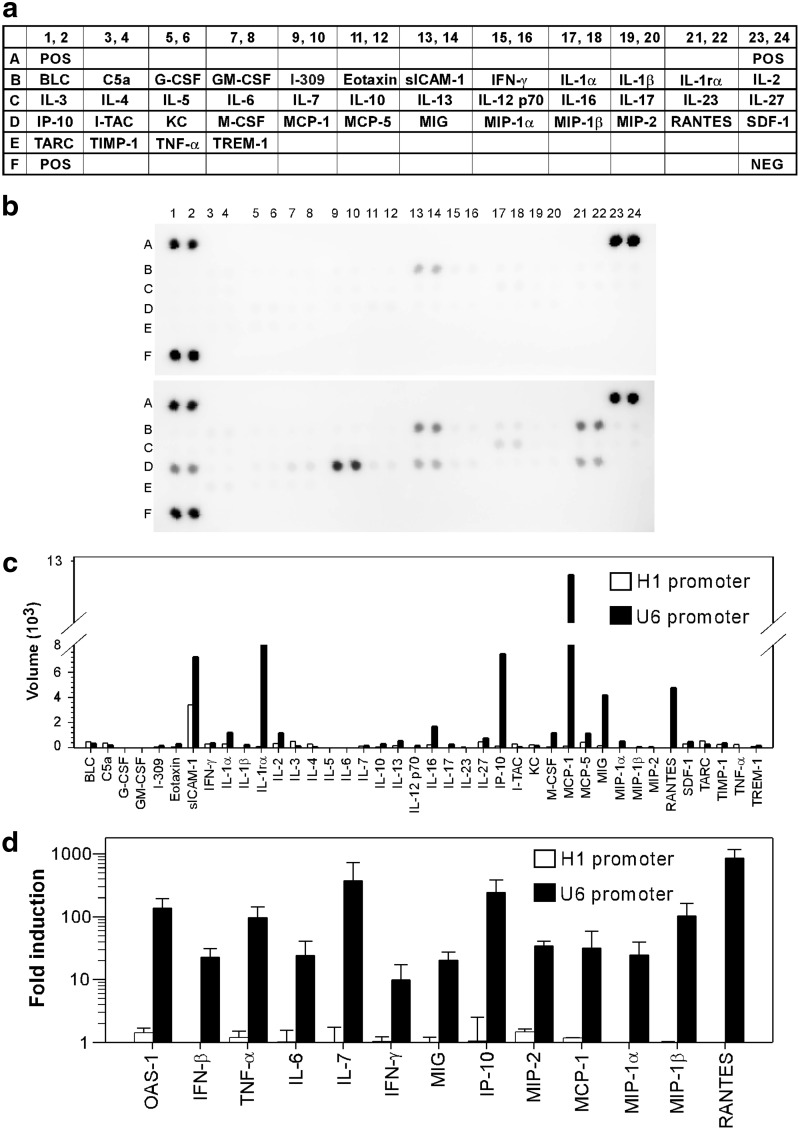
Similar articles
-
Use of RNA interference to modulate liver adenoma development in a murine model transgenic for hepatitis B virus.Gene Ther. 2012 Jan;19(1):25-33. doi: 10.1038/gt.2011.60. Epub 2011 May 12. Gene Ther. 2012. PMID: 21562593
-
Comparative study of anti-hepatitis B virus RNA interference by double-stranded adeno-associated virus serotypes 7, 8, and 9.Mol Ther. 2009 Feb;17(2):352-9. doi: 10.1038/mt.2008.245. Epub 2008 Dec 9. Mol Ther. 2009. PMID: 19066602 Free PMC article.
-
Effective inhibition of HBV replication in vivo by anti-HBx short hairpin RNAs.Mol Ther. 2006 Feb;13(2):411-21. doi: 10.1016/j.ymthe.2005.10.013. Epub 2005 Dec 5. Mol Ther. 2006. PMID: 16337206
-
Expression of shRNA from a tissue-specific pol II promoter is an effective and safe RNAi therapeutic.Mol Ther. 2008 Sep;16(9):1630-6. doi: 10.1038/mt.2008.144. Epub 2008 Jul 29. Mol Ther. 2008. PMID: 18665161
-
The short hairpin RNA driven by polymerase II suppresses both wild-type and lamivudine-resistant hepatitis B virus strains.Antivir Ther. 2007;12(6):865-76. Antivir Ther. 2007. PMID: 17926641
Cited by
-
In vivo genome editing in animals using AAV-CRISPR system: applications to translational research of human disease.F1000Res. 2017 Dec 20;6:2153. doi: 10.12688/f1000research.11243.1. eCollection 2017. F1000Res. 2017. PMID: 29333255 Free PMC article. Review.
-
Adipose Tissue: An Emerging Target for Adeno-associated Viral Vectors.Mol Ther Methods Clin Dev. 2020 Sep 20;19:236-249. doi: 10.1016/j.omtm.2020.09.009. eCollection 2020 Dec 11. Mol Ther Methods Clin Dev. 2020. PMID: 33102616 Free PMC article. Review.
-
Progress and Prospects of Anti-HBV Gene Therapy Development.Int J Mol Sci. 2015 Jul 31;16(8):17589-610. doi: 10.3390/ijms160817589. Int J Mol Sci. 2015. PMID: 26263978 Free PMC article. Review.
-
Inhibition of hepatitis B virus replication by helper dependent adenoviral vectors expressing artificial anti-HBV pri-miRs from a liver-specific promoter.Biomed Res Int. 2014;2014:718743. doi: 10.1155/2014/718743. Epub 2014 Jun 5. Biomed Res Int. 2014. PMID: 25003129 Free PMC article.
-
Thyroid hormone suppresses hepatocarcinogenesis via DAPK2 and SQSTM1-dependent selective autophagy.Autophagy. 2016 Dec;12(12):2271-2285. doi: 10.1080/15548627.2016.1230583. Epub 2016 Sep 21. Autophagy. 2016. PMID: 27653365 Free PMC article.
References
-
- Bridge A.J. Pebernard S. Ducraux A., et al. Induction of an interferon response by RNAi vectors in mammalian cells. Nat. Genet. 2003;34:263–264. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
