

Structural evidence: a single charged residue affects substrate binding in cytochrome P450 BM-3
- PMID: 23829560
- PMCID: PMC5945292
- DOI: 10.1021/bi4000645
Structural evidence: a single charged residue affects substrate binding in cytochrome P450 BM-3
Abstract
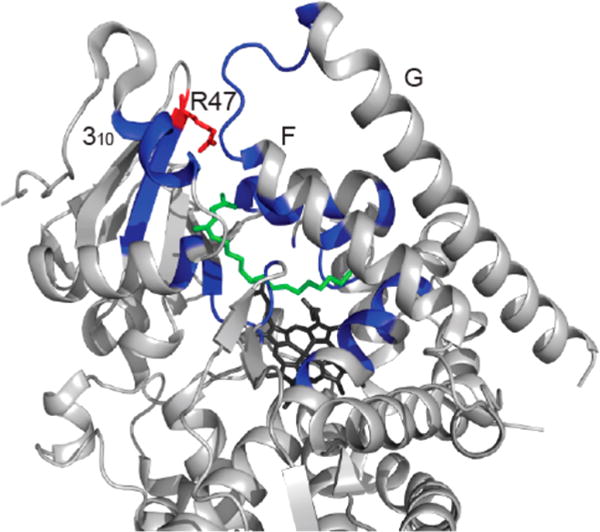
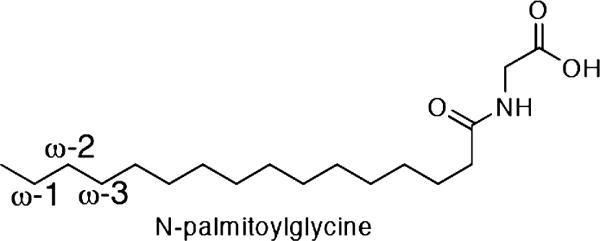
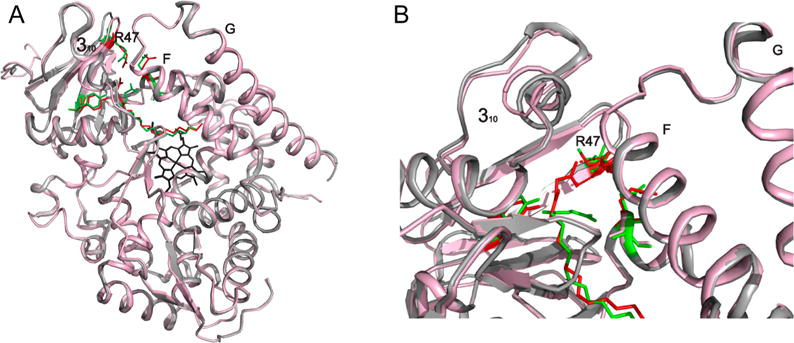
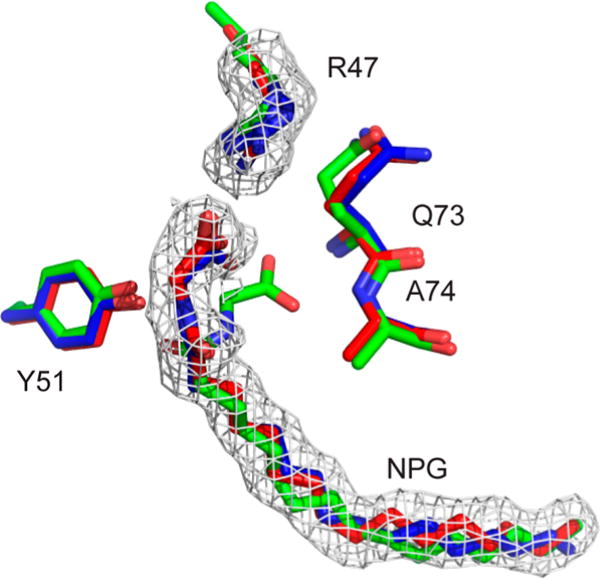
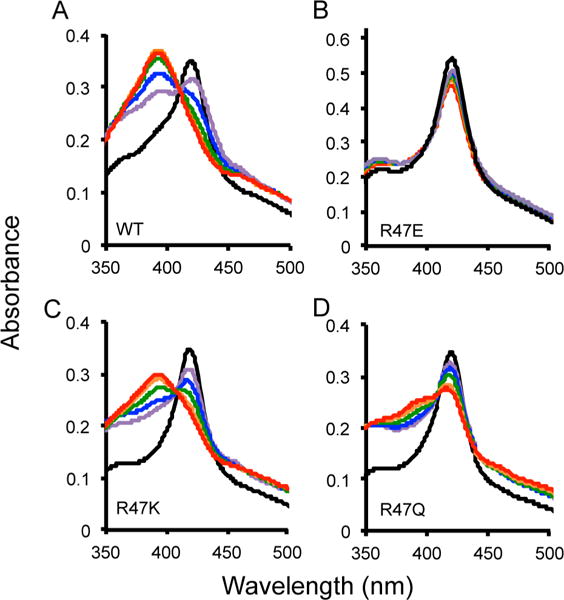
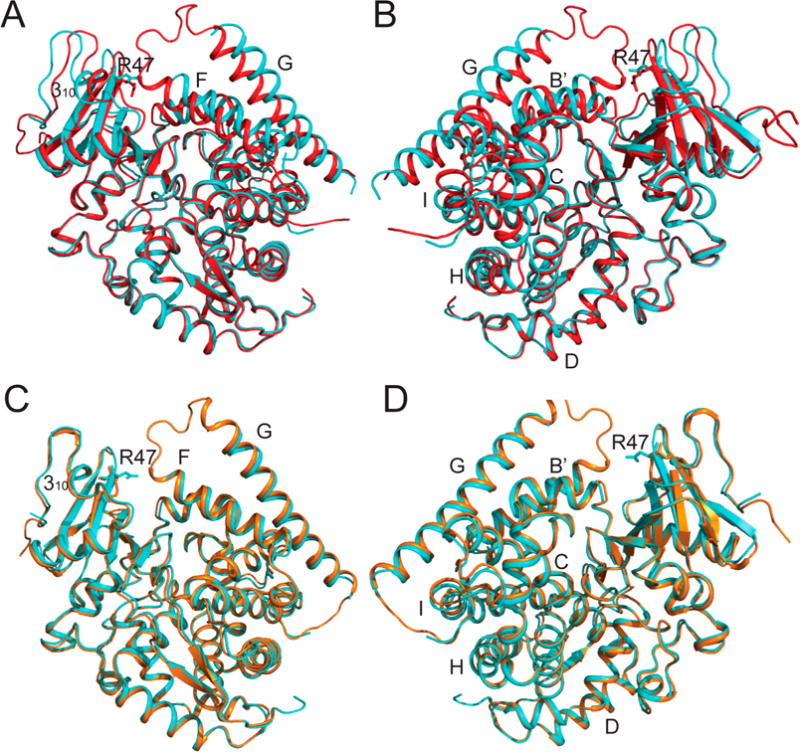
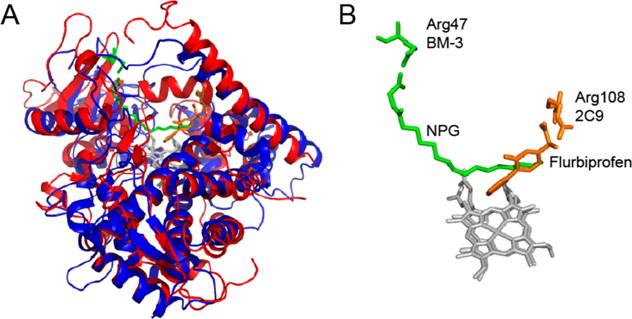
Cytochrome P450 BM-3 is a bacterial enzyme with sequence similarity to mammalian P450s that catalyzes the hydroxylation of fatty acids with high efficiency. Enzyme-substrate binding and dynamics has been an important topic of study for cytochromes P450 because most of the crystal structures of substrate-bound structures show the complex in an inactive state. We have determined a new crystal structure for cytochrome P450 BM-3 in complex with N-palmitoylglycine (NPG), which unexpectedly showed a direct bidentate ion pair between NPG and arginine 47 (R47). We further explored the role of R47, the only charged residue in the binding pocket in cytochrome P450 BM-3, through mutagenesis and crystallographic studies. The mutations of R47 to glutamine (R47Q), glutamic acid (R47E), and lysine (R47K) were designed to investigate the role of its charge in binding and catalysis. The oppositely charged R47E mutation had the greatest effect on activity and binding. The crystal structure of R47E BMP shows that the glutamic acid side chain is blocking the entrance to the binding pocket, accounting for NPG's low binding affinity and charge repulsion. For R47Q and R47K BM-3, the mutations caused only a slight change in kcat and a large change in Km and Kd, which suggests that R47 mostly is involved in binding and that our crystal structure, 4KPA , represents an initial binding step in the P450 cycle.
Conflict of interest statement
The authors declare no competing financial interest.
Figures
Similar articles
-
Filling a hole in cytochrome P450 BM3 improves substrate binding and catalytic efficiency.J Mol Biol. 2007 Oct 26;373(3):633-51. doi: 10.1016/j.jmb.2007.08.015. Epub 2007 Aug 21. J Mol Biol. 2007. PMID: 17868686
-
Structural basis of steroid binding and oxidation by the cytochrome P450 CYP109E1 from Bacillus megaterium.FEBS J. 2016 Nov;283(22):4128-4148. doi: 10.1111/febs.13911. Epub 2016 Oct 17. FEBS J. 2016. PMID: 27686671 Free PMC article.
-
Interactions of substrates at the surface of P450s can greatly enhance substrate potency.Biochemistry. 2007 Dec 11;46(49):14010-7. doi: 10.1021/bi701667m. Epub 2007 Nov 16. Biochemistry. 2007. PMID: 18004886
-
Fatty acid metabolism, conformational change, and electron transfer in cytochrome P-450(BM-3).Biochim Biophys Acta. 1999 Nov 23;1441(2-3):141-9. doi: 10.1016/s1388-1981(99)00161-4. Biochim Biophys Acta. 1999. PMID: 10570242 Review.
-
Structure, dynamics, and function of the monooxygenase P450 BM-3: insights from computer simulations studies.J Phys Condens Matter. 2015 Jul 15;27(27):273102. doi: 10.1088/0953-8984/27/27/273102. Epub 2015 Jun 10. J Phys Condens Matter. 2015. PMID: 26061496 Review.
Cited by
-
Structure-Function Analysis of the Self-Sufficient CYP102 Family Provides New Insights into Their Biochemistry.Int J Mol Sci. 2025 Feb 28;26(5):2161. doi: 10.3390/ijms26052161. Int J Mol Sci. 2025. PMID: 40076785 Free PMC article.
-
Insights into an efficient light-driven hybrid P450 BM3 enzyme from crystallographic, spectroscopic and biochemical studies.Biochim Biophys Acta. 2016 Dec;1864(12):1732-1738. doi: 10.1016/j.bbapap.2016.09.005. Epub 2016 Sep 14. Biochim Biophys Acta. 2016. PMID: 27639964 Free PMC article.
-
The use of reaction timecourses to determine the level of minor contaminants in enzyme preparations.Anal Biochem. 2014 Apr 1;450:20-6. doi: 10.1016/j.ab.2013.12.031. Epub 2014 Jan 3. Anal Biochem. 2014. PMID: 24393696 Free PMC article.
References
-
- Schoch GA, Yano JK, Wester MR, Griffin KJ, Stout CD, Johnson EF. Structure of human microsomal cytochrome P4502C8 - Evidence for a peripheral fatty acid binding site. J Biol Chem. 2004;279:9497–9503. - PubMed
-
- Williams PA, Cosme J, Vinkovic DM, Ward A, Angove HC, Day PJ, Vonrhein C, Tickle IJ, Jhoti H. Crystal structures of human cytochrome P450 3A4 bound to metyrapone and progesterone. Science. 2004;305:683–686. - PubMed
-
- Williams PA, Cosme J, Ward A, Angova HC, Vinkovic DM, Jhoti H. Crystal structure of human cytochrome P4502C9 with bound warfarin. Nature. 2003;424:464–468. - PubMed
-
- Wester MR, Yano JK, Schoch GA, Yang C, Griffin KJ, Stout CD, Johnson EF. The structure of human cytochrome P4502C9 complexed with flurbiprofen at 2.0-angstrom resolution. J Biol Chem. 2004;279:35630–35637. - PubMed
-
- Narhi LO, Fulco AJ. Phenobarbital induction of a soluble cytochrome P-450-dependent fatty acid monooxygenase in Bacillus megaterium. J Biol Chem. 1982;257:2147–2150. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
