

Continuous DOPA synthesis from a single AAV: dosing and efficacy in models of Parkinson's disease
- PMID: 23831692
- PMCID: PMC3703607
- DOI: 10.1038/srep02157
Continuous DOPA synthesis from a single AAV: dosing and efficacy in models of Parkinson's disease
Abstract
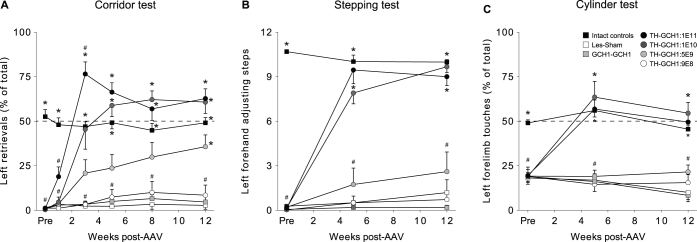
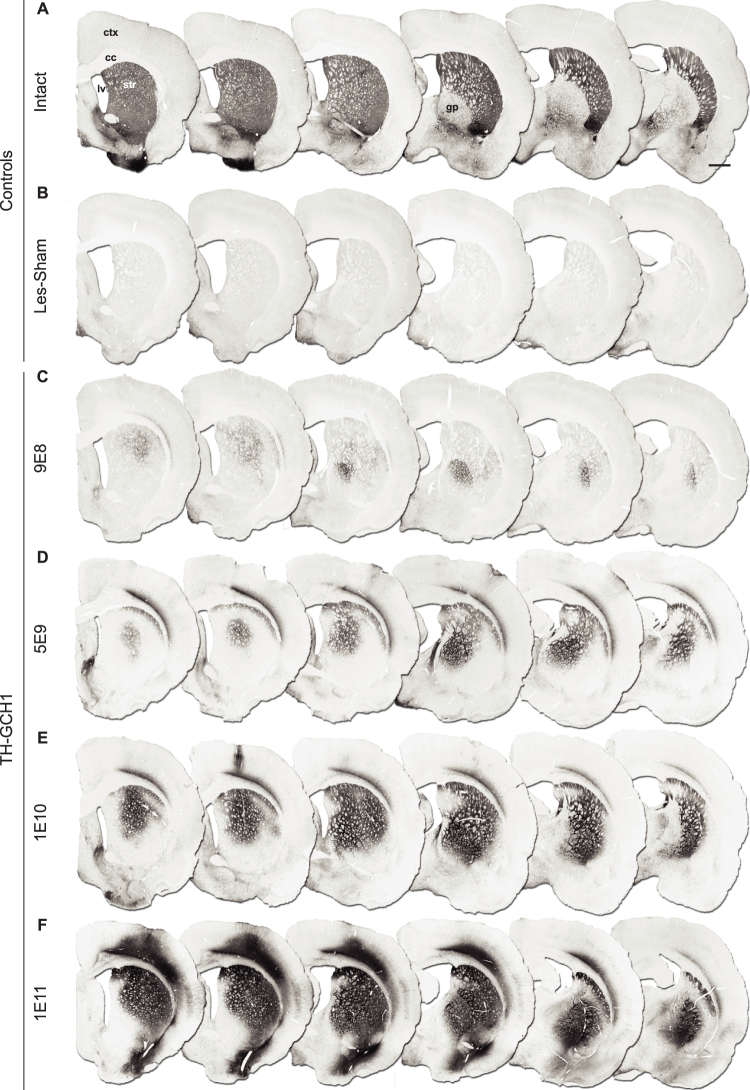
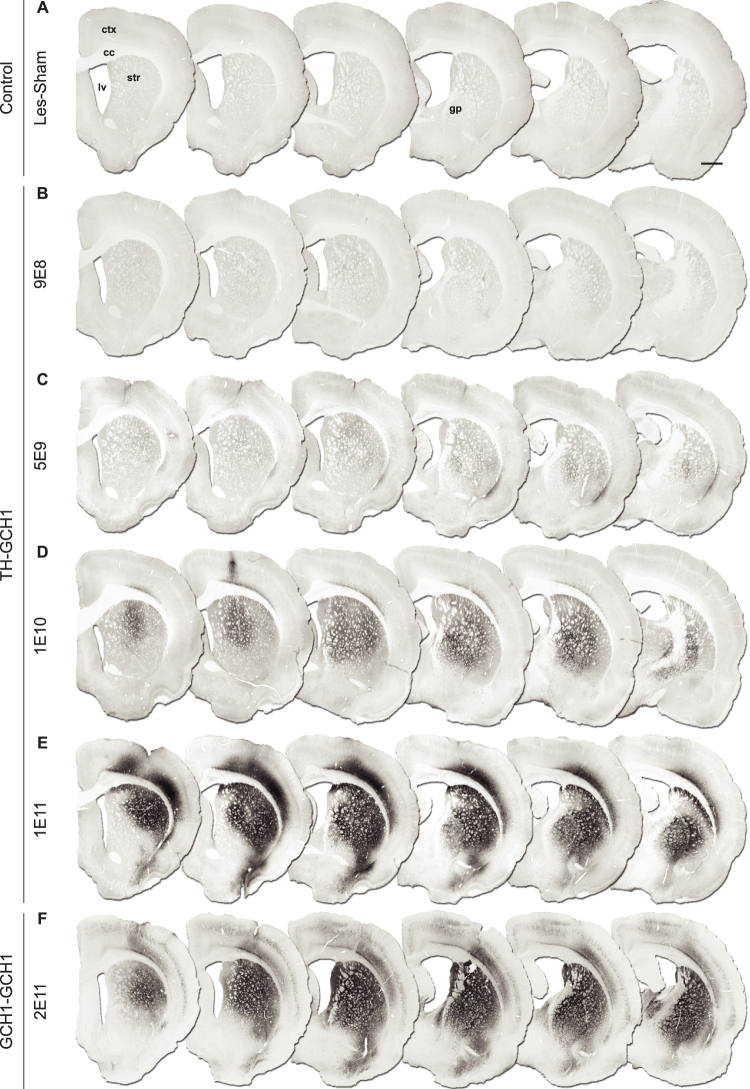
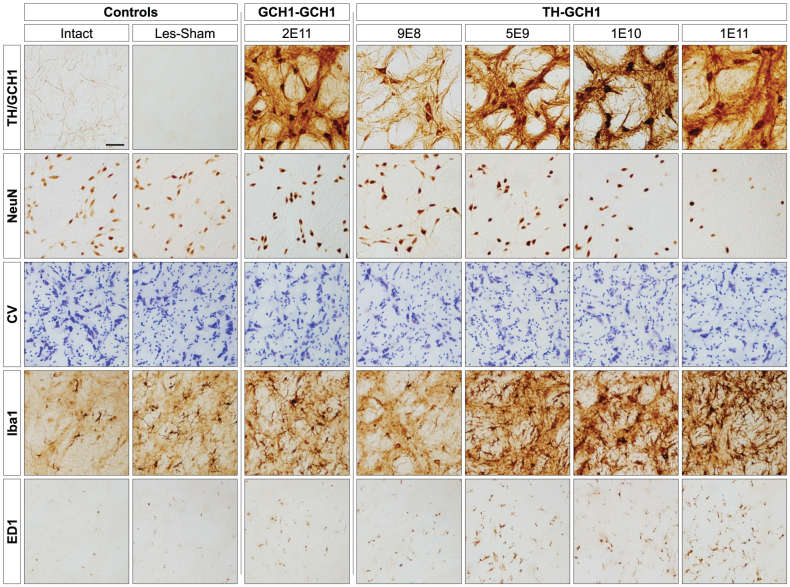
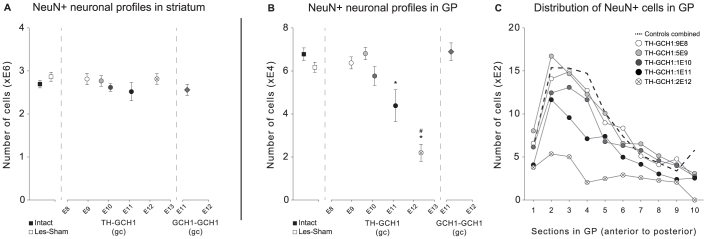
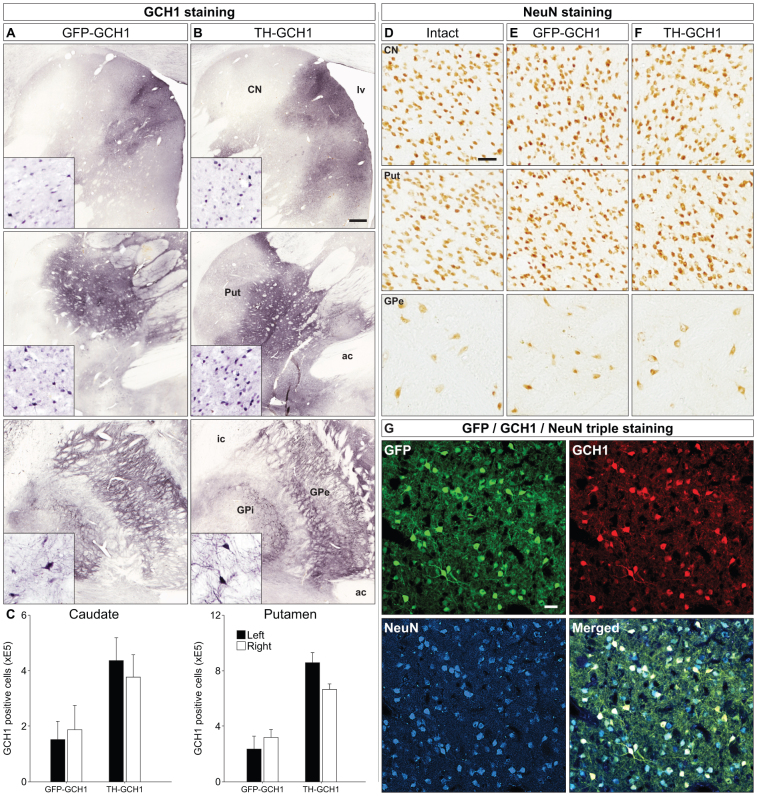
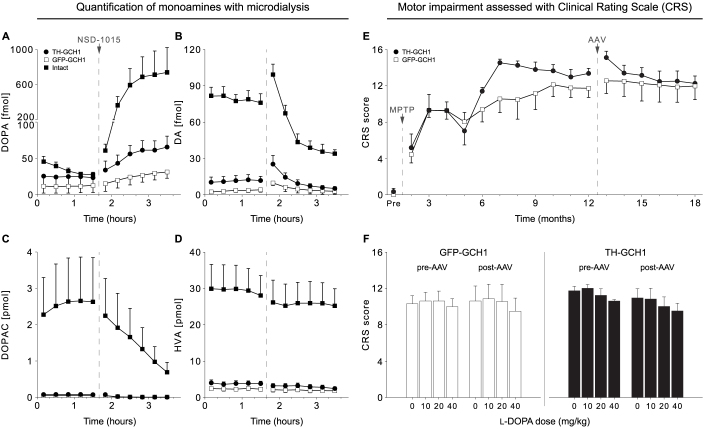
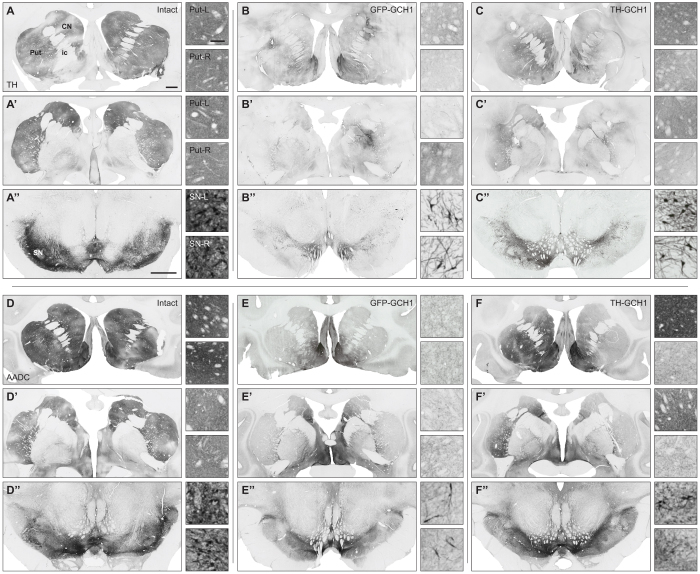
We used a single adeno-associated viral (AAV) vector co-expressing tyrosine hydroxylase (TH) and GTP cyclohydrolase 1 (GCH1) to investigate the relationship between vector dose, and the magnitude and rate of recovery in hemi-parkinsonian rats. Intrastriatal injections of >1E10 genomic copies (gc) of TH-GCH1 vector resulted in complete recovery in drug-naïve behavior tests. Lower vector dose gave partial to no functional improvement. Stereological quantification revealed no striatal NeuN+ cell loss in any of the groups, whereas a TH-GCH1 dose of >1E11 gc resulted in cell loss in globus pallidus. Thus, a TH-GCH1 dose of 1E10 gc gave complete recovery without causing neuronal loss. Safety and efficacy was also studied in non-human primates where the control vector resulted in co-expression of the transgenes in caudate-putamen. In the TH-GCH1 group, GCH1 expression was robust but TH was not detectable. Moreover, TH-GCH1 treatment did not result in functional improvement in non-human primates.
Conflict of interest statement
DK and TB are co-inventors of a patent application on the construct reported in this paper. TB is a founding-director of a company that intends to develop this for commercial purposes. DK receives financial compensation in the form of retainer fees, milestones and royalty payments. JHK is a Founding Scientist and currently an SAB member of Ceregene Inc. EC, NN, GS, EN and YC have no conflict of interest.
Figures

References
-
- Fahn S. Parkinson disease, the effect of levodopa, and the ELLDOPA trial. Earlier vs Later L-DOPA. Arch Neurol 56, 529–535 (1999). - PubMed
-
- Nutt J. G. & Holford N. H. The response to levodopa in Parkinson's disease: imposing pharmacological law and order. Ann Neurol 39, 561–573 (1996). - PubMed
-
- Obeso J. A., Olanow C. W. & Nutt J. G. Levodopa motor complications in Parkinson's disease. Trends Neurosci 23, S2–7 (2000). - PubMed
-
- Kurlan R. et al. Duodenal delivery of levodopa for on-off fluctuations in parkinsonism: preliminary observations. Ann Neurol 20, 262–265 (1986). - PubMed
-
- Nilsson D. et al. Long-term intraduodenal infusion of a water based levodopa-carbidopa dispersion in very advanced Parkinson's disease. Acta Neurol Scand 97, 175–183 (1998). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
