

Plasticity of renal erythropoietin-producing cells governs fibrosis
- PMID: 23833259
- PMCID: PMC3785278
- DOI: 10.1681/ASN.2013010030
Plasticity of renal erythropoietin-producing cells governs fibrosis
Abstract
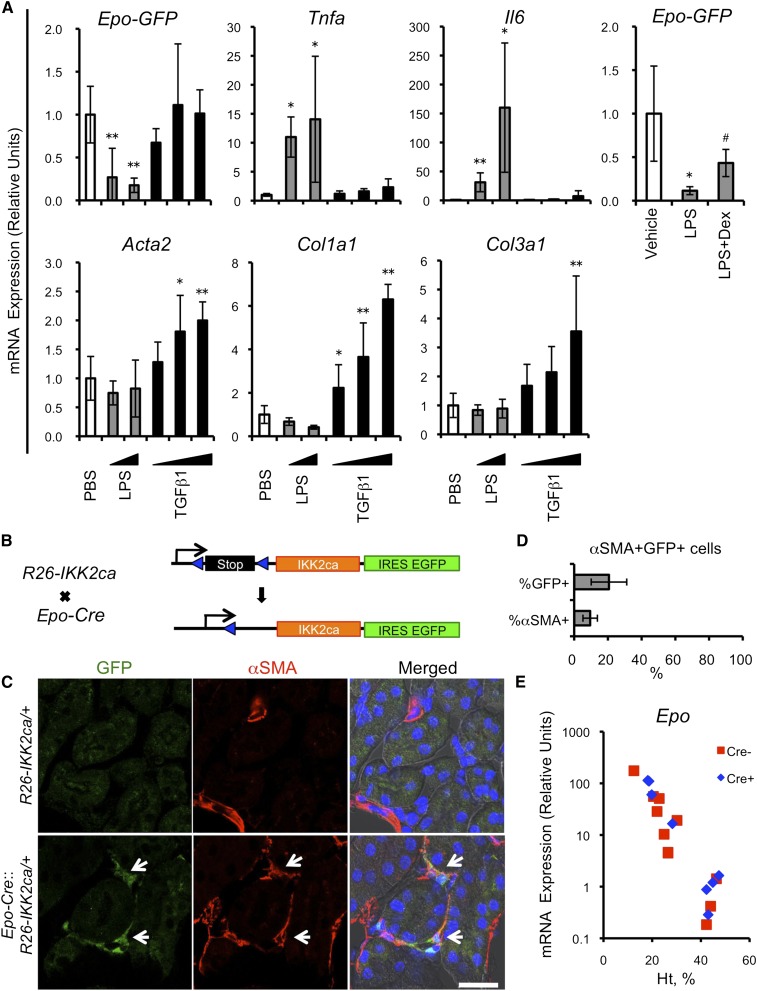
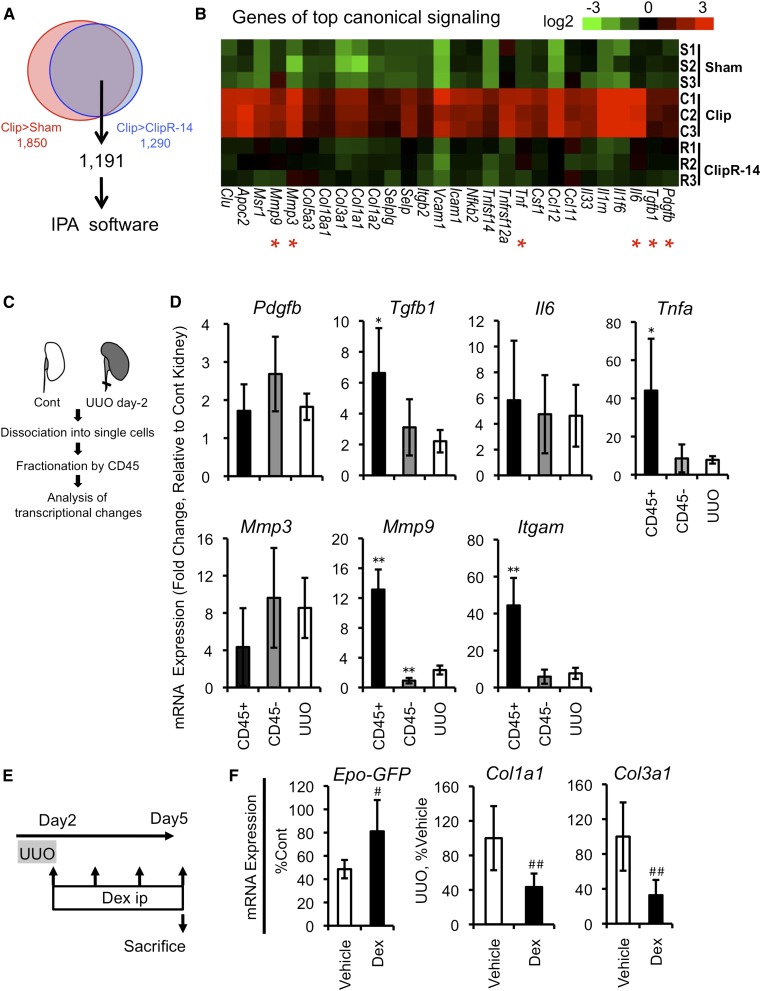
CKD progresses with fibrosis and erythropoietin (Epo)-dependent anemia, leading to increased cardiovascular complications, but the mechanisms linking Epo-dependent anemia and fibrosis remain unclear. Here, we show that the cellular phenotype of renal Epo-producing cells (REPs) alternates between a physiologic Epo-producing state and a pathologic fibrogenic state in response to microenvironmental signals. In a novel mouse model, unilateral ureteral obstruction-induced inflammatory milieu activated NFκB and Smad signaling pathways in REPs, rapidly repressed the Epo-producing potential of REPs, and led to myofibroblast transformation of these cells. Moreover, we developed a unique Cre-based cell-fate tracing method that marked current and/or previous Epo-producing cells and revealed that the majority of myofibroblasts are derived from REPs. Genetic induction of NFκB activity selectively in REPs resulted in myofibroblastic transformation, indicating that NFκB signaling elicits a phenotypic switch. Reversing the unilateral ureteral obstruction-induced inflammatory microenvironment restored the Epo-producing potential and the physiologic phenotype of REPs. This phenotypic reversion was accelerated by anti-inflammatory therapy. These findings demonstrate that REPs possess cellular plasticity, and suggest that the phenotypic transition of REPs to myofibroblasts, modulated by inflammatory molecules, underlies the connection between anemia and renal fibrosis in CKD.
Figures







Comment in
-
Chronic kidney disease: Fibrosis and anaemia in CKD--two beasts, one ancestor.Nat Rev Nephrol. 2013 Oct;9(10):563-5. doi: 10.1038/nrneph.2013.179. Epub 2013 Sep 3. Nat Rev Nephrol. 2013. PMID: 23999397 No abstract available.
References
-
- Maxwell PH, Osmond MK, Pugh CW, Heryet A, Nicholls LG, Tan CC, Doe BG, Ferguson DJP, Johnson MH, Ratcliffe PJ: Identification of the renal erythropoietin-producing cells using transgenic mice. Kidney Int 44: 1149–1162, 1993 - PubMed
-
- Obara N, Suzuki N, Kim K, Nagasawa T, Imagawa S, Yamamoto M: Repression via the GATA box is essential for tissue-specific erythropoietin gene expression. Blood 111: 5223–5232, 2008 - PubMed
-
- Wenger RH, Hoogewijs D: Regulated oxygen sensing by protein hydroxylation in renal erythropoietin-producing cells. Am J Physiol Renal Physiol 298: F1287–F1296, 2010 - PubMed
-
- Eckardt KU, Koury ST, Tan CC, Schuster SJ, Kaissling B, Ratcliffe PJ, Kurtz A: Distribution of erythropoietin producing cells in rat kidneys during hypoxic hypoxia. Kidney Int 43: 815–823, 1993 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
