

tRNA tKUUU, tQUUG, and tEUUC wobble position modifications fine-tune protein translation by promoting ribosome A-site binding
- PMID: 23836657
- PMCID: PMC3725067
- DOI: 10.1073/pnas.1300781110
tRNA tKUUU, tQUUG, and tEUUC wobble position modifications fine-tune protein translation by promoting ribosome A-site binding
Abstract
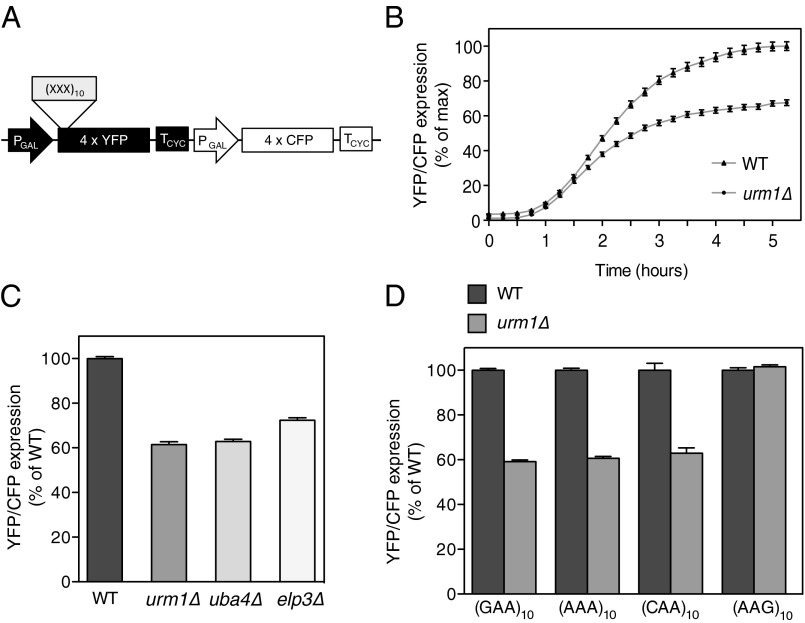
tRNA modifications are crucial to ensure translation efficiency and fidelity. In eukaryotes, the URM1 and ELP pathways increase cellular resistance to various stress conditions, such as nutrient starvation and oxidative agents, by promoting thiolation and methoxycarbonylmethylation, respectively, of the wobble uridine of cytoplasmic (tK(UUU)), (tQ(UUG)), and (tE(UUC)). Although in vitro experiments have implicated these tRNA modifications in modulating wobbling capacity and translation efficiency, their exact in vivo biological roles remain largely unexplored. Using a combination of quantitative proteomics and codon-specific translation reporters, we find that translation of a specific gene subset enriched for AAA, CAA, and GAA codons is impaired in the absence of URM1- and ELP-dependent tRNA modifications. Moreover, in vitro experiments using native tRNAs demonstrate that both modifications enhance binding of tK(UUU) to the ribosomal A-site. Taken together, our data suggest that tRNA thiolation and methoxycarbonylmethylation regulate translation of genes with specific codon content.
Keywords: SILAC; systems biology; translation regulation.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Agris PF, Vendeix FA, Graham WD. tRNA’s wobble decoding of the genome: 40 years of modification. J Mol Biol. 2007;366(1):1–13. - PubMed
-
- Krüger MK, Pedersen S, Hagervall TG, Sørensen MA. The modification of the wobble base of tRNAGlu modulates the translation rate of glutamic acid codons in vivo. J Mol Biol. 1998;284(3):621–631. - PubMed
-
- Murphy FV, 4th, Ramakrishnan V, Malkiewicz A, Agris PF. The role of modifications in codon discrimination by tRNA(Lys)UUU. Nat Struct Mol Biol. 2004;11(12):1186–1191. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
