

Review
doi: 10.1083/jcb.201304099.
Big steps toward understanding dynein
Affiliations
- PMID: 23836927
- PMCID: PMC3704981
- DOI: 10.1083/jcb.201304099
Item in Clipboard
Review
Big steps toward understanding dynein
J Cell Biol.
.
Abstract
Dynein is a microtubule-based molecular motor that is involved in various biological functions, such as axonal transport, mitosis, and cilia/flagella movement. Although dynein was discovered 50 years ago, the progress of dynein research has been slow due to its large size and flexible structure. Recent progress in understanding the force-generating mechanism of dynein using x-ray crystallography, cryo-electron microscopy, and single molecule studies has provided key insight into the structure and mechanism of action of this complex motor protein.
Figures
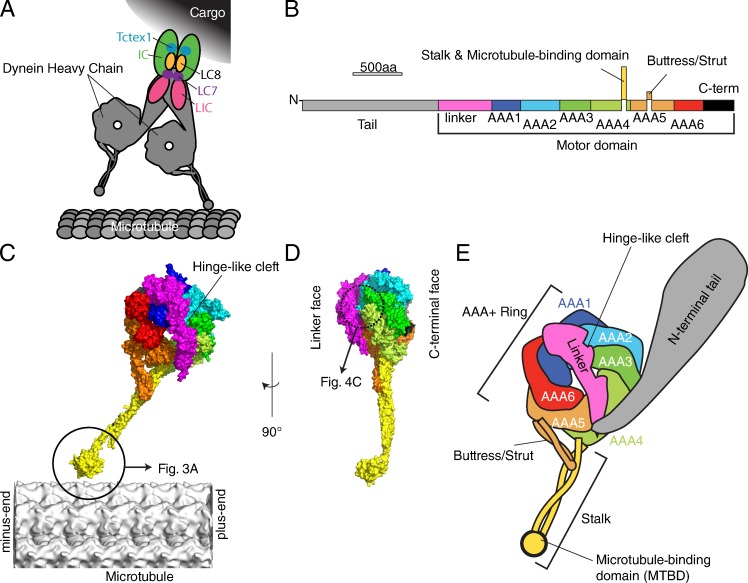
Atomic structures of cytoplasmic dynein. (A) Schematic structure of cytoplasmic dynein complex, adapted from Allan (2011). (B) The primary structure of cytoplasmic dynein. (C and D) The atomic model of D. discoideum cytoplasmic dynein motor domain (PDB accession no. 3VKG) overlaid on a microtubule (EMDB-5193; Sui and Downing, 2010) according to the orientation determined by Mizuno et al. (2007) (C) Side view. (D) View from the plus end of microtubule. (E) Schematic domain structure of dynein.
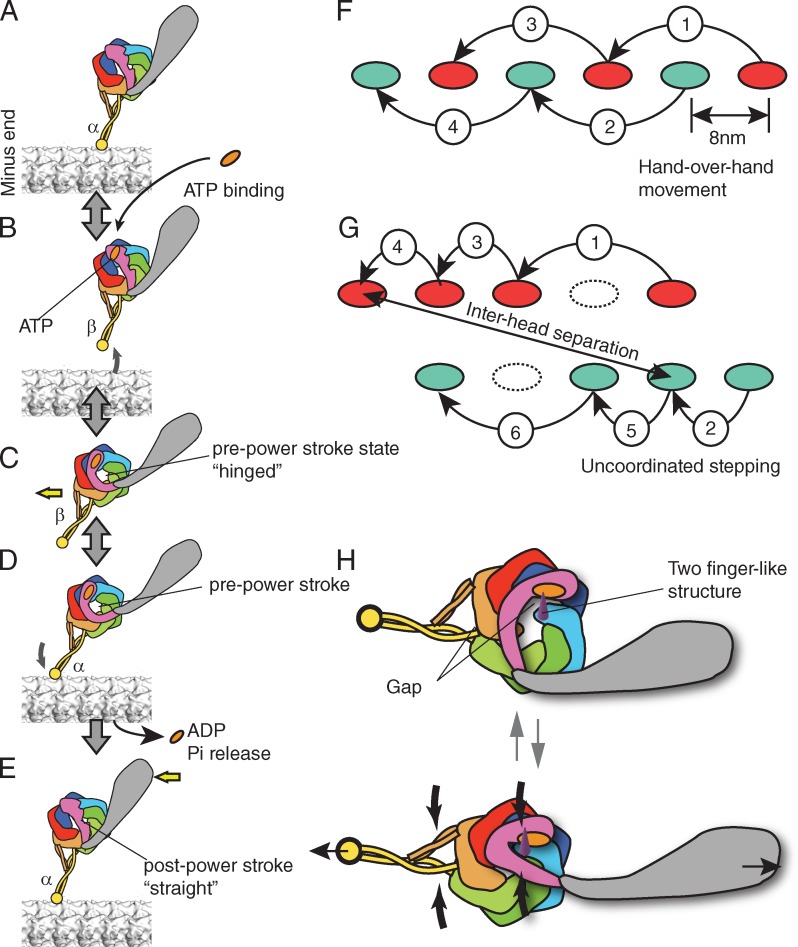
Presumed chemomechanical cycle and stepping of dynein. (A–E) Chemomechanical cycle of dynein. The pre- and post-power stroke states are also called the primed and unprimed states, respectively. The registries of the stalk coiled coil are denoted as α and β according to Gibbons et al. (2005). (F and G) Processive movement of kinesin (F) and dynein (G). (F) Hand-over-hand movement of kinesin. A step by one head (red) is always followed by the step of another head (green). The stepping of kinesin is on one protofilament of microtubule. (G) Presumed stepping of dynein. The step size varies and the interhead separation can be large. A step by one head (red) is not always flowed by the step of another head (green). (H) A model of strain-based dynein ATPase activation. (G, top) Without strain, the gap between the AAA1 and AAA2 is open and the motor domain cannot hydrolyze ATP. (G, bottom) Under a strain imposed between MTBD and tail (thin black arrows), the gap becomes smaller (thick black arrows) and turns on ATP hydrolysis by dynein.
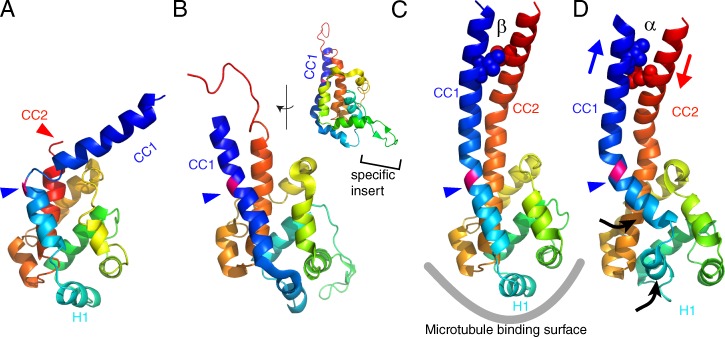
Atomic models of the MTBD of dynein. (A) D. discoideum cytoplasmic dynein (PDB accession no. 3VKH). (B) C. reinhardtii dynein c (PDB accession no. 2RR7). The inset shows the side view, highlighting the dynein c–specific insert. (C) Mouse cytoplasmic dynein (PDB accession no. 3ERR). (D) Mouse cytoplasmic dynein fit to the MTBD–microtubule complex derived from cryo-EM (PDB accession no. 3J1T). All the MTBD structures were aligned using least square fits and color-coded with a gradient from the N to C terminus. CC1, coiled coil helix 1; CC2, coiled coil helix 2. The blue arrowheads points to the junction between MTBD and the stalk, where a well-conserved proline residue (colored pink) is located. In C and D, two residues (isoleucine 3269 and leucine 3417) are shown as spheres. The two residues form hydrophobic contacts in the β-registry (C), whereas they are separated in the α-registry (D) because of the sliding between the two α-helices (blue and red arrows). Conformational changes observed in the mouse dynein MTBD in complex with a microtubule by cryo-EM are shown by black arrows. Note that the cryo-EM density map does not have enough resolution to observe sliding between CC1 and CC2. The sliding was modeled based on targeted molecular dynamics (Redwine et al., 2012).
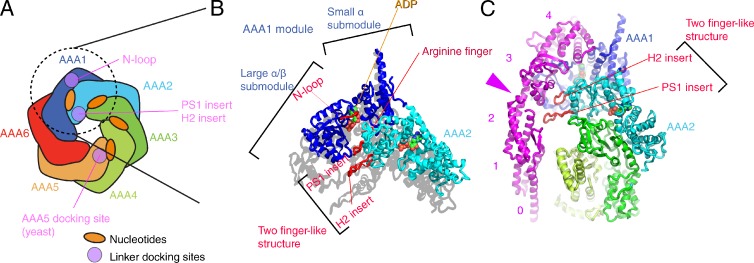
Structures around the first ATP binding site. (A) Schematic domain structure of the head domain. Regions contacting the linker domain are colored purple. (B) AAA submodules surrounding the first nucleotide-binding pocket (PDB accession no. 3VKG, chain A). The linker is connected to AAA1 domain by the “N-loop.” To highlight that the two finger-like structures are protruding, the shadow of the atomic structure has been cast on the plane parallel to the head domain. (C) Interaction between the linker and the two finger-like structures. The pink arrowhead points to the hinge-like structure of the linker. The pink numbers indicates the subdomain of the linker.
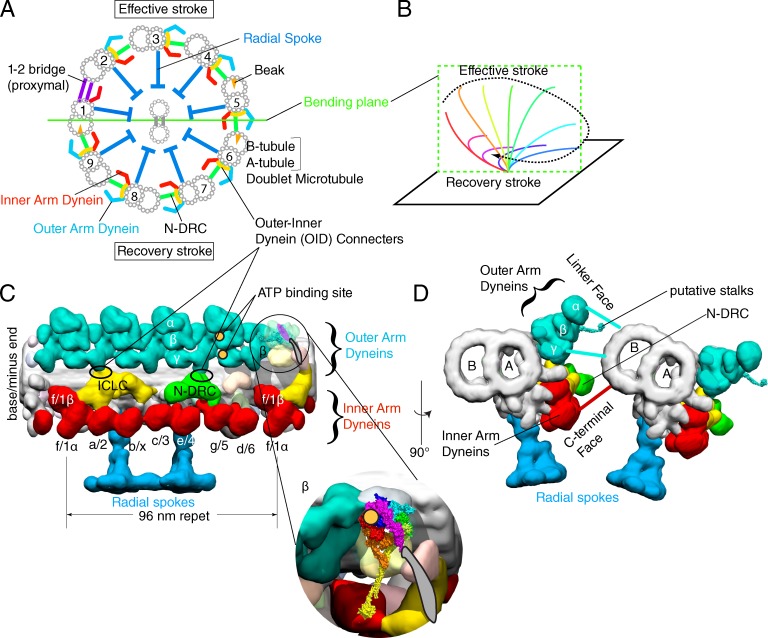
Arrangement of axonemal dyneins. (A) The schematic structure of the motile 9 + 2 axoneme, viewed from the base of flagella. (B) Quasi-planar asymmetric movement of the 9 + 2 axoneme typically observed in trachea cilia or in C. reinhardtii flagella. (C and D) 3D structure of a 96-nm repeat of doublet microtubules in the distal/central region of C. reinhardtii flagella (EMDB-2132; Bui et al., 2012). N-DRC, the nexin-dynein regulatory complex; ICLC, intermediate chain/light chain complex. Inner arm dynein subspecies are labeled according to Bui et al. (2012) and Lin et al. (2012). To avoid the confusion with the linker domain of dynein, the structures connecting between outer and inner arm dyneins are labeled as “connecters,” which are normally called “linkers.” Putative ATP binding sites of outer arm dynein determined by biotin-ADP (Oda et al., 2013) are indicated by orange circles. The atomic structure of cytoplasmic dynein is placed into the β-heavy chain of outer arm dynein and its enlarged view is shown in the inset. (D) Two doublet microtubules, viewed from the base of flagella.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
