

Translesion DNA polymerases
- PMID: 23838442
- PMCID: PMC3783050
- DOI: 10.1101/cshperspect.a010363
Translesion DNA polymerases
Abstract
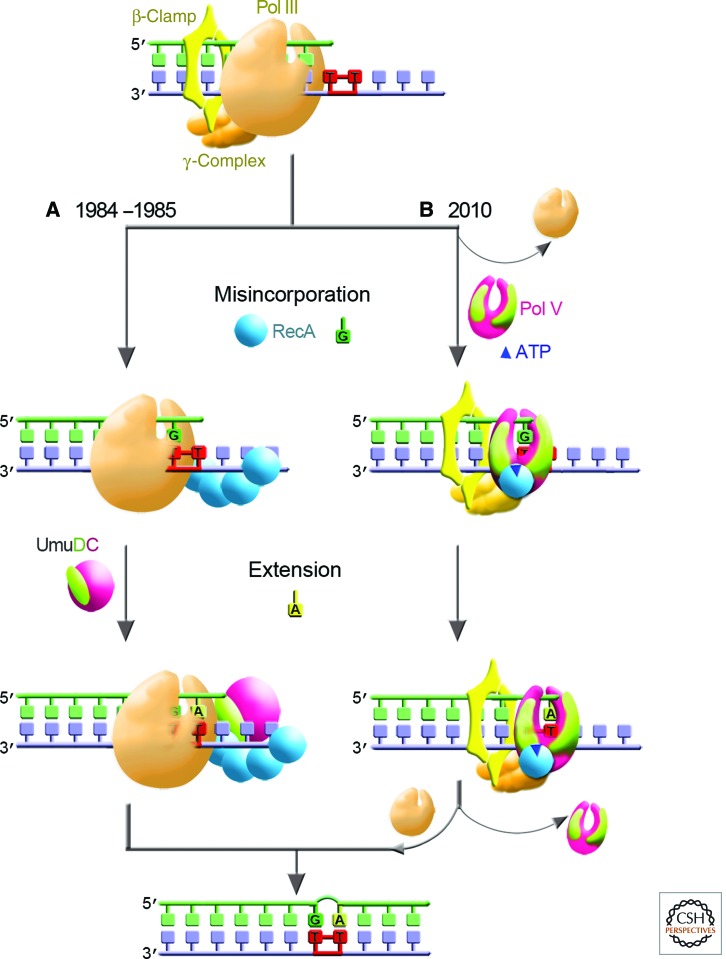
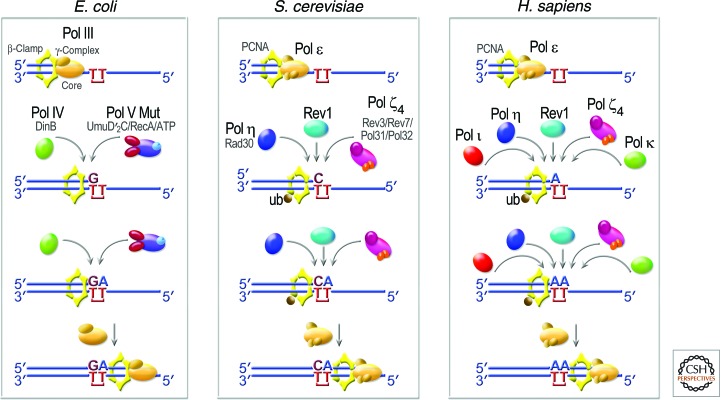
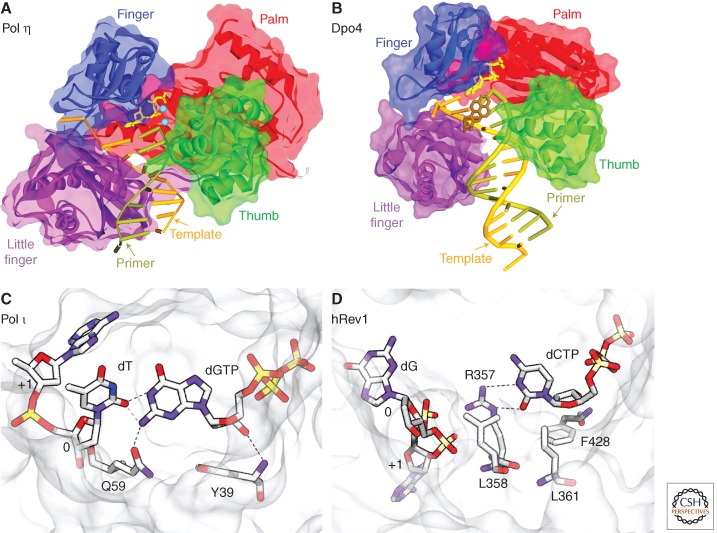
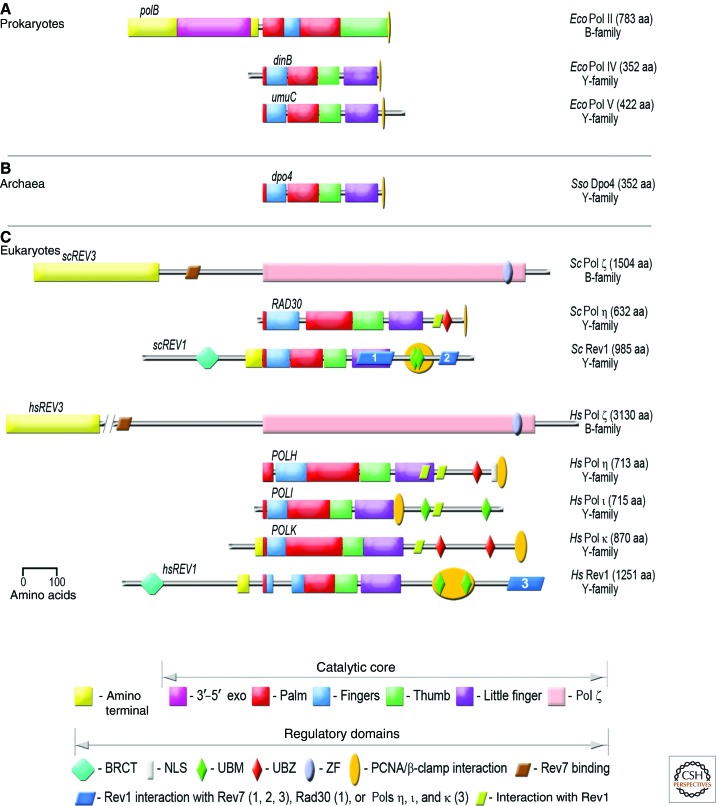
Living cells are continually exposed to DNA-damaging agents that threaten their genomic integrity. Although DNA repair processes rapidly target the damaged DNA for repair, some lesions nevertheless persist and block genome duplication by the cell's replicase. To avoid the deleterious consequence of a stalled replication fork, cells use specialized polymerases to traverse the damage. This process, termed "translesion DNA synthesis" (TLS), affords the cell additional time to repair the damage before the replicase returns to complete genome duplication. In many cases, this damage-tolerance mechanism is error-prone, and cell survival is often associated with an increased risk of mutagenesis and carcinogenesis. Despite being tightly regulated by a variety of transcriptional and posttranslational controls, the low-fidelity TLS polymerases also gain access to undamaged DNA where their inaccurate synthesis may actually be beneficial for genetic diversity and evolutionary fitness.
Figures
References
-
- Anderson WF, Prince DB, Yu H, McEntee K, Goodman MF 1994. Crystallization of DNA polymerase II from Escherichia coli. J Mol Biol 238: 120–122 - PubMed
-
- Beaber JW, Hochhut B, Waldor MK 2004. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature 427: 72–74 - PubMed
-
- Bebenek K, Kunkel TA 2004. Functions of DNA polymerases. Adv Protein Chem 69: 137–165 - PubMed
-
- Becherel OJ, Fuchs RP, Wagner J 2002. Pivotal role of the β-clamp in translesion DNA synthesis and mutagenesis in E. coli cells. DNA Repair 1: 703–708 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases