

Intrinsic selectivity of Notch 1 for Delta-like 4 over Delta-like 1
- PMID: 23839946
- PMCID: PMC3757209
- DOI: 10.1074/jbc.M113.454850
Intrinsic selectivity of Notch 1 for Delta-like 4 over Delta-like 1
Abstract
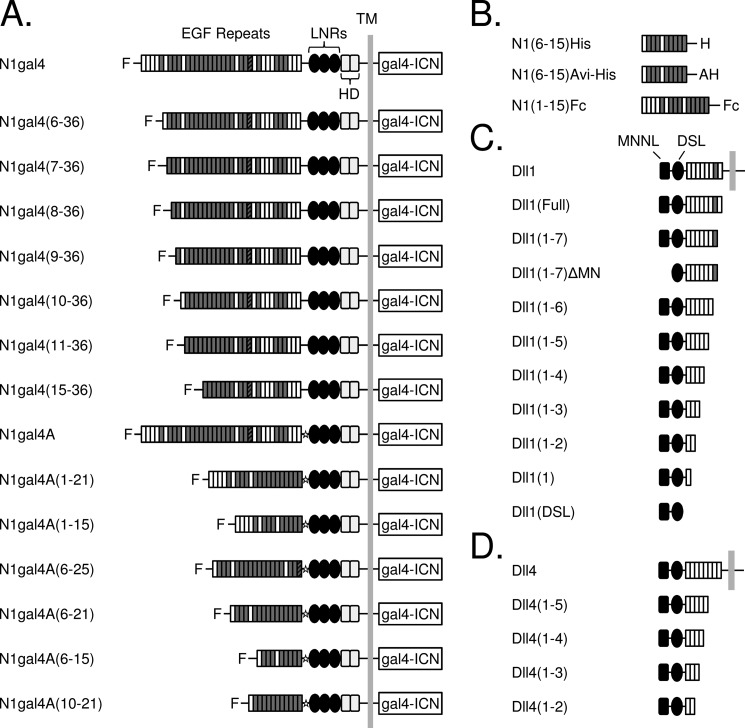
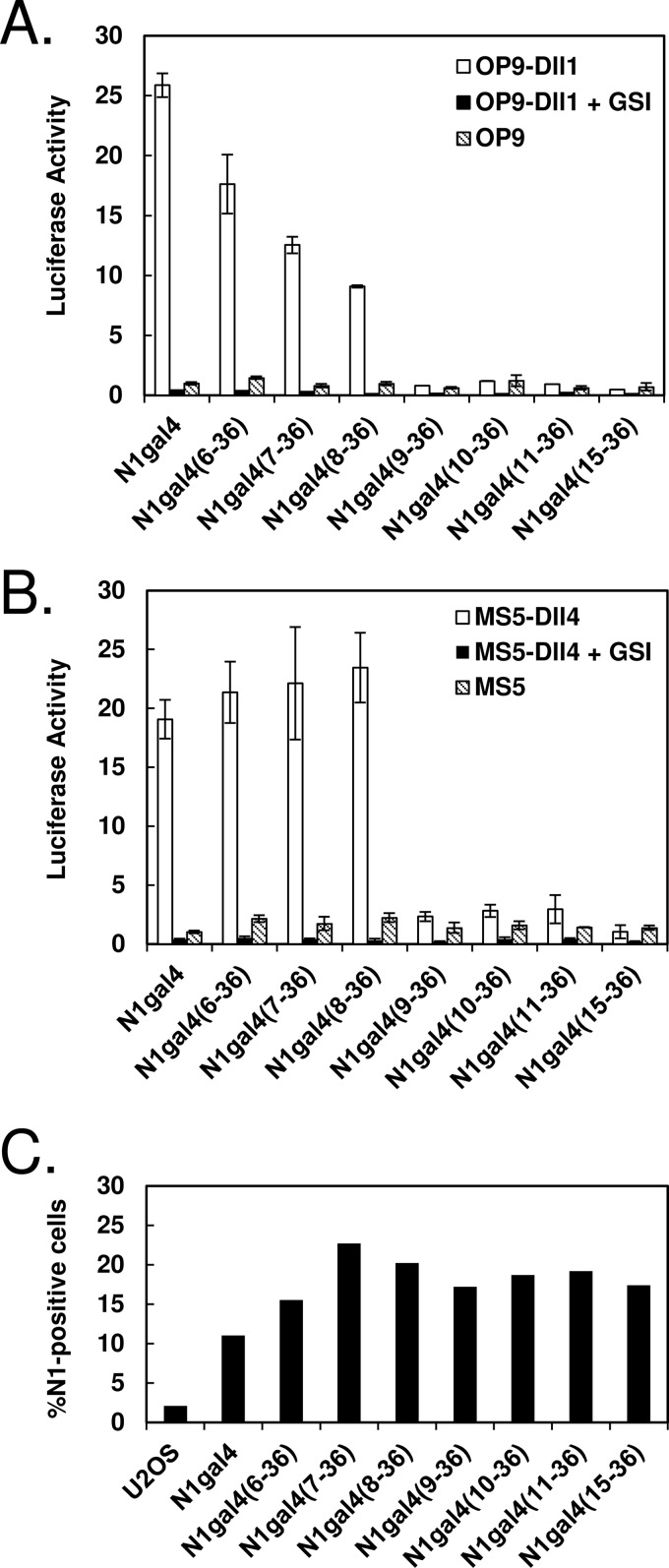
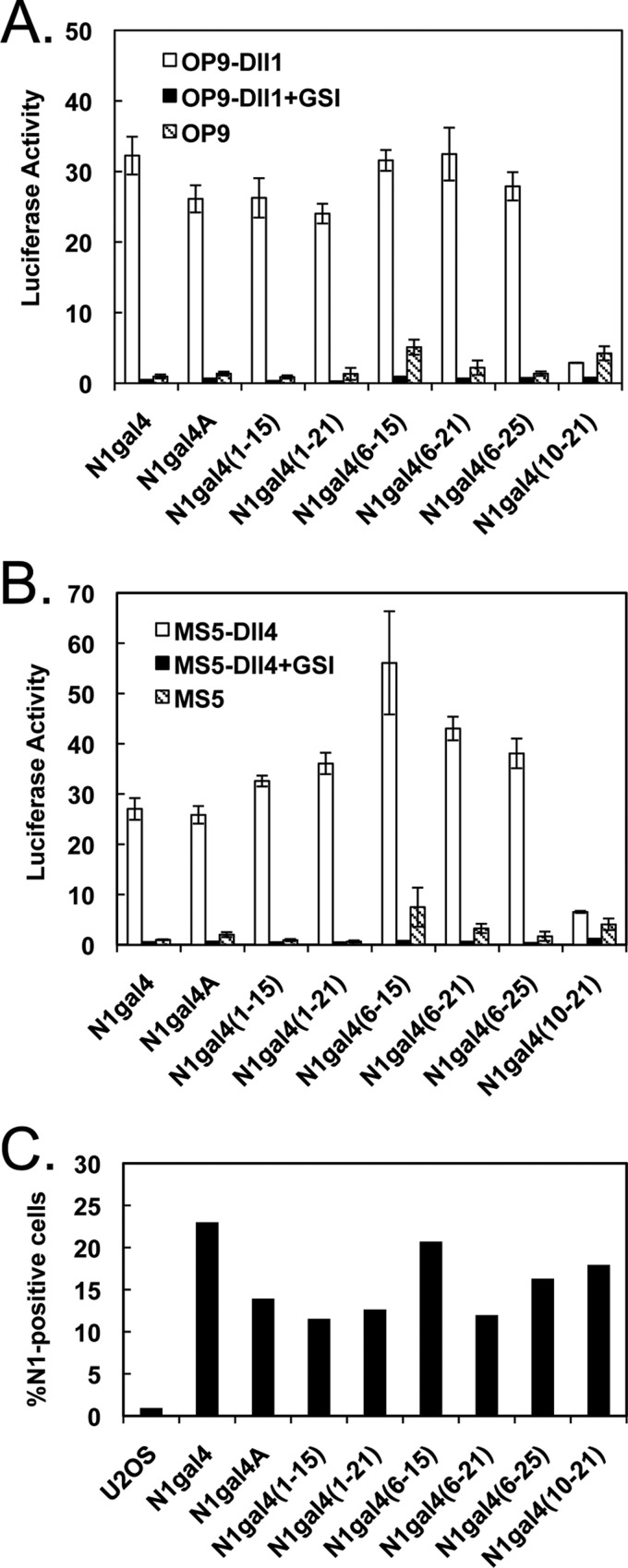
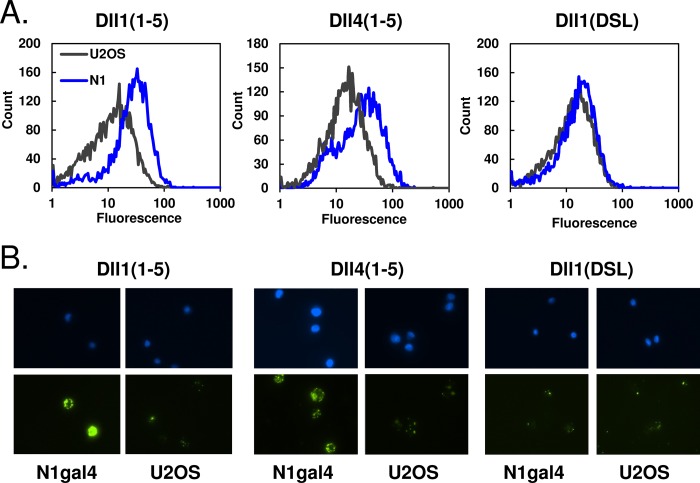
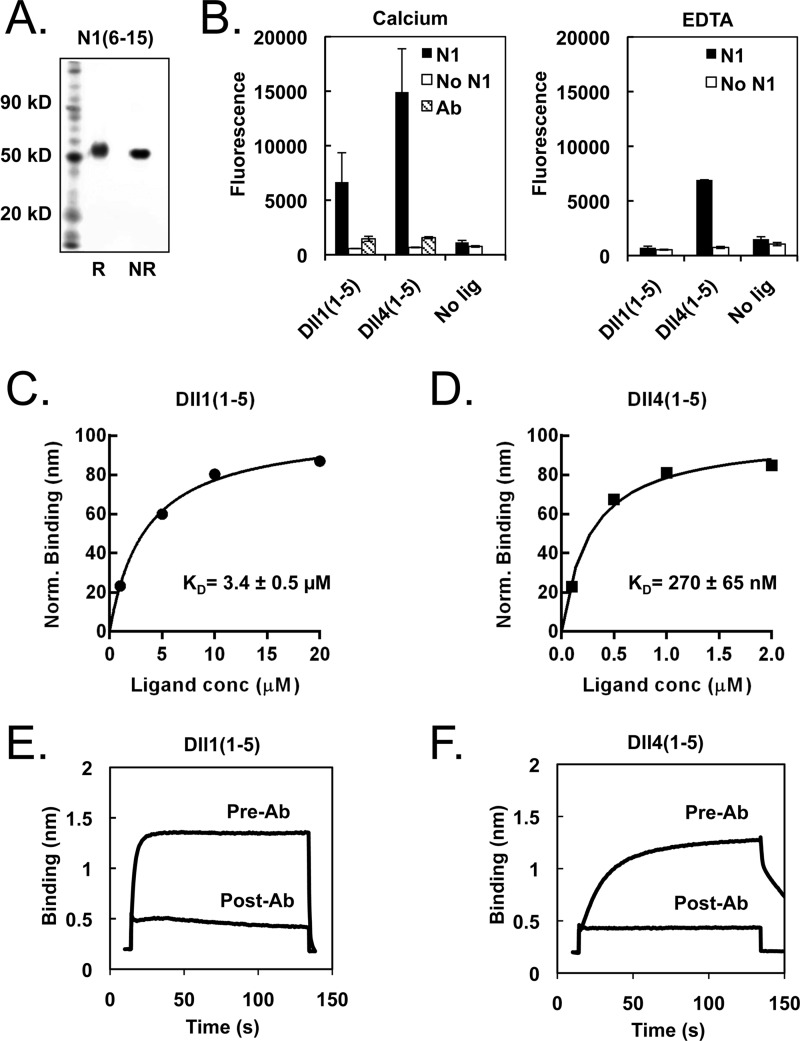
Notch signaling makes critical contributions to cell fate determination in all metazoan organisms, yet remarkably little is known about the binding affinity of the four mammalian Notch receptors for their three Delta-like and two Jagged family ligands. Here, we utilized signaling assays and biochemical studies of purified recombinant ligand and receptor molecules to investigate the differences in signaling behavior and intrinsic affinity between Notch1-Dll1 and Notch1-Dll4 complexes. Systematic deletion mutagenesis of the human Notch1 ectodomain revealed that epidermal growth factor (EGF) repeats 6-15 are sufficient to maintain signaling in a reporter assay at levels comparable with the full-length receptor, and identified important contributions from EGF repeats 8-10 in conveying an activating signal in response to either Dll1 or Dll4. Truncation studies of the Dll1 and Dll4 ectodomains showed that the MNNL-EGF3 region was both necessary and sufficient for full activation. Plate-based and cell binding assays revealed a specific, calcium-dependent interaction between cell-surface and recombinant Notch receptors and ligand molecules. Finally, direct measurement of the binding affinity of Notch1 EGF repeats 6-15 for Dll1 and Dll4 revealed that Dll4 binds with at least an order of magnitude higher affinity than Dll1. Together, these studies give new insights into the features of ligand recognition by Notch1, and highlight how intrinsic differences in the biochemical behavior of receptor-ligand complexes can influence receptor-mediated responses of developmental signaling pathways.
Keywords: Biolayer Interferometry; Cell Signaling; Cell Surface Receptor; Cell-Cell Interaction; Notch Pathway; Receptor Structure-Function; Receptor-Ligand Interaction; Recombinant Protein Expression.
Figures

References
-
- Artavanis-Tsakonas S., Rand M. D., Lake R. J. (1999) Notch signaling. Cell fate control and signal integration in development. Science 284, 770–776 - PubMed
-
- Bray S. J. (2006) Notch signalling. A simple pathway becomes complex. Nat. Rev. Mol. Cell Biol. 7, 678–689 - PubMed
-
- Fukuda D., Aikawa E., Swirski F. K., Novobrantseva T. I., Kotelianski V., Gorgun C. Z., Chudnovskiy A., Yamazaki H., Croce K., Weissleder R., Aster J. C., Hotamisligil G. S., Yagita H., Aikawa M. (2012) Notch ligand Delta-like 4 blockade attenuates atherosclerosis and metabolic disorders. Proc. Natl. Acad. Sci. U.S.A. 109, E1868–E1877 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
