

A Low Temperature Limit for Life on Earth
- PMID: 23840425
- PMCID: PMC3686811
- DOI: 10.1371/journal.pone.0066207
A Low Temperature Limit for Life on Earth
Abstract
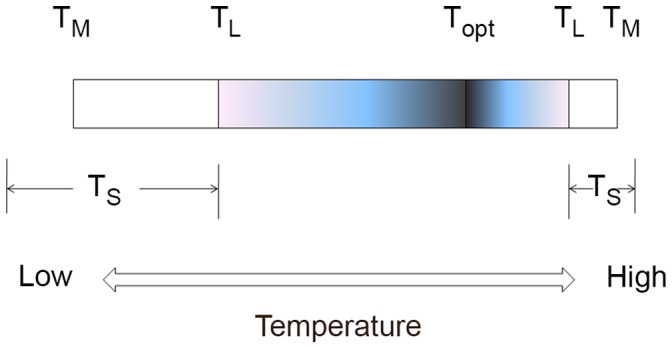
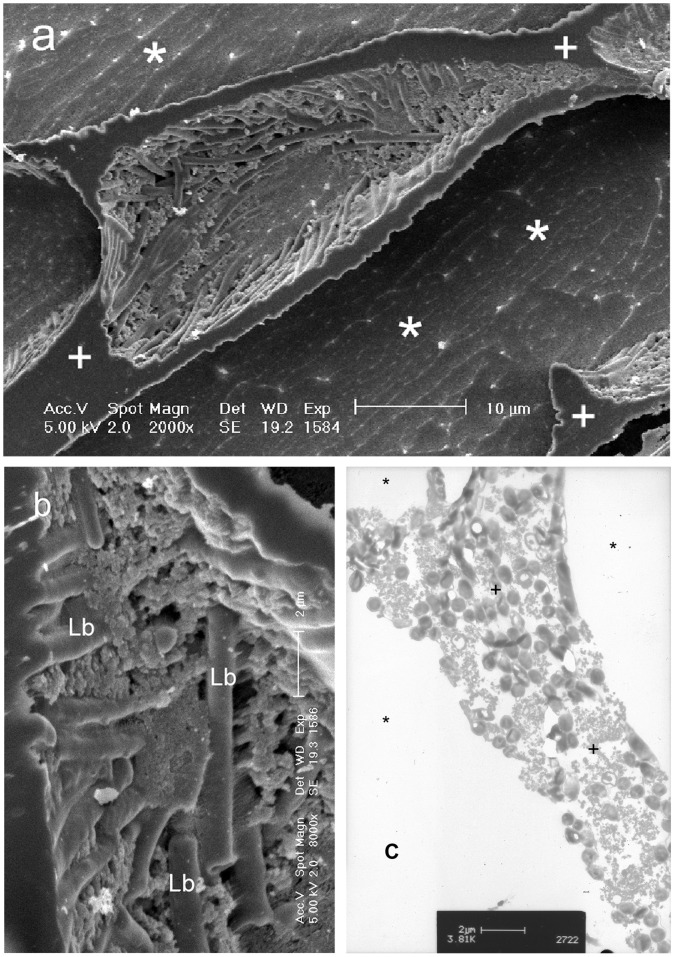
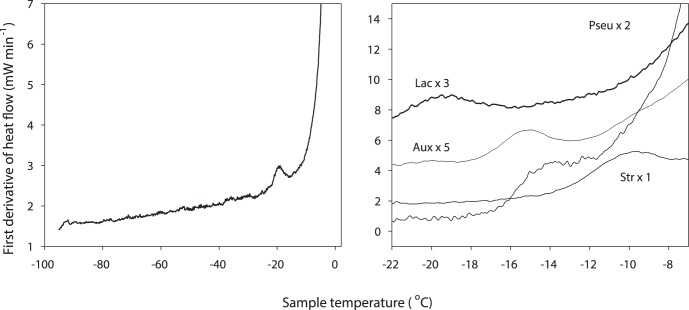
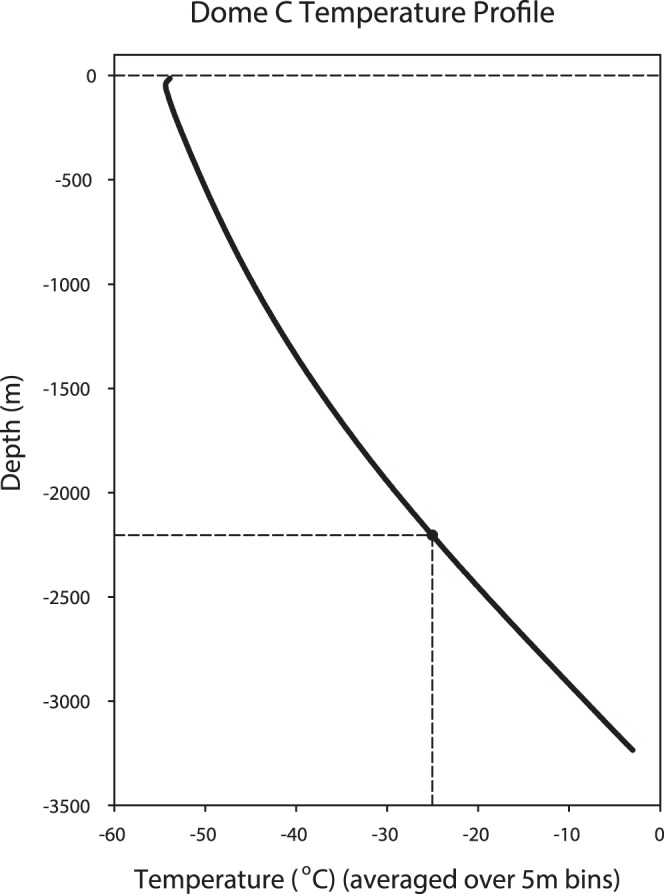
There is no generally accepted value for the lower temperature limit for life on Earth. We present empirical evidence that free-living microbial cells cooling in the presence of external ice will undergo freeze-induced desiccation and a glass transition (vitrification) at a temperature between -10°C and -26°C. In contrast to intracellular freezing, vitrification does not result in death and cells may survive very low temperatures once vitrified. The high internal viscosity following vitrification means that diffusion of oxygen and metabolites is slowed to such an extent that cellular metabolism ceases. The temperature range for intracellular vitrification makes this a process of fundamental ecological significance for free-living microbes. It is only where extracellular ice is not present that cells can continue to metabolise below these temperatures, and water droplets in clouds provide an important example of such a habitat. In multicellular organisms the cells are isolated from ice in the environment, and the major factor dictating how they respond to low temperature is the physical state of the extracellular fluid. Where this fluid freezes, then the cells will dehydrate and vitrify in a manner analogous to free-living microbes. Where the extracellular fluid undercools then cells can continue to metabolise, albeit slowly, to temperatures below the vitrification temperature of free-living microbes. Evidence suggests that these cells do also eventually vitrify, but at lower temperatures that may be below -50°C. Since cells must return to a fluid state to resume metabolism and complete their life cycle, and ice is almost universally present in environments at sub-zero temperatures, we propose that the vitrification temperature represents a general lower thermal limit to life on Earth, though its precise value differs between unicellular (typically above -20°C) and multicellular organisms (typically below -20°C). Few multicellular organisms can, however, complete their life cycle at temperatures below ∼-2°C.
Conflict of interest statement
Figures
References
-
- Kashefi K, Lovley DR (2003) Extending the upper temperature limit for life. Science 301(5635): 934–934. - PubMed
-
- Kerry KR, Grace DR, Williams R, Burton HR (1977) Studies on some saline lakes of the Vestfold Hills, Antarctica. In: Llano GA, editor. Adaptations within Antarctic ecosystems. Washington, D.C.: Smithsonian Institution. 839–858.
-
- Tominaga H, Fukui F (1981) Saline lakes in Syowa Oasis, Antarctica. Hydrobiologia 82(3): 375–389.
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
