

Igf Signaling is Required for Cardiomyocyte Proliferation during Zebrafish Heart Development and Regeneration
- PMID: 23840646
- PMCID: PMC3694143
- DOI: 10.1371/journal.pone.0067266
Igf Signaling is Required for Cardiomyocyte Proliferation during Zebrafish Heart Development and Regeneration
Abstract
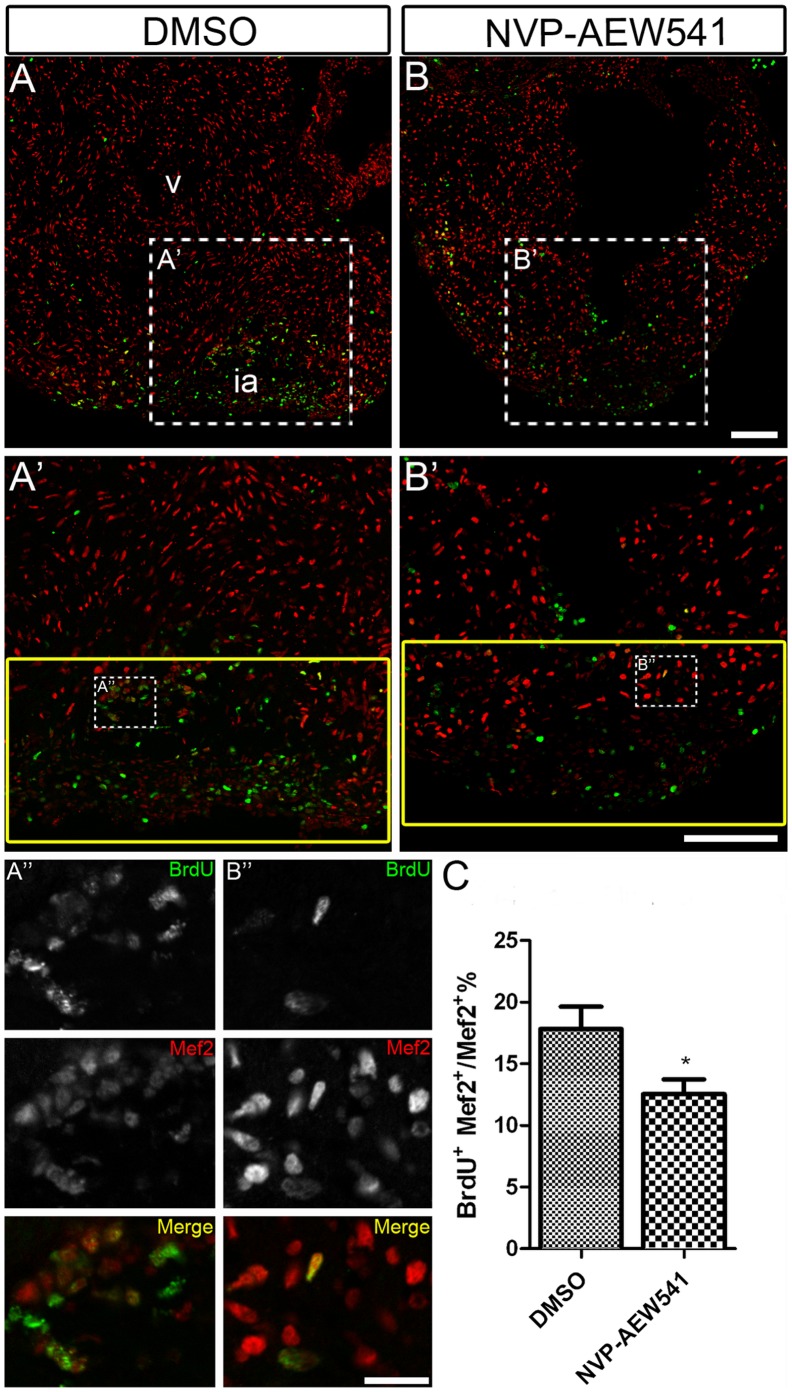
Unlike its mammalian counterpart, the adult zebrafish heart is able to fully regenerate after severe injury. One of the most important events during the regeneration process is cardiomyocyte proliferation, which results in the replacement of lost myocardium. Growth factors that induce cardiomyocyte proliferation during zebrafish heart regeneration remain to be identified. Signaling pathways important for heart development might be reutilized during heart regeneration. IGF2 was recently shown to be important for cardiomyocyte proliferation and heart growth during mid-gestation heart development in mice, although its role in heart regeneration is unknown. We found that expression of igf2b was upregulated during zebrafish heart regeneration. Following resection of the ventricle apex, igf2b expression was detected in the wound, endocardium and epicardium at a time that coincides with cardiomyocyte proliferation. Transgenic zebrafish embryos expressing a dominant negative form of Igf1 receptor (dn-Igf1r) had fewer cardiomyocytes and impaired heart development, as did embryos treated with an Igf1r inhibitor. Moreover, inhibition of Igf1r signaling blocked cardiomyocyte proliferation during heart development and regeneration. We found that Igf signaling is required for a subpopulation of cardiomyocytes marked by gata4:EGFP to contribute to the regenerating area. Our findings suggest that Igf signaling is important for heart development and myocardial regeneration in zebrafish.
Conflict of interest statement
Figures






References
-
- Li F, Wang X, Capasso JM, Gerdes AM (1996) Rapid transition of cardiac myocytes from hyperplasia to hypertrophy during postnatal development. J Mol Cell Cardiol 28: 1737–1746. - PubMed
-
- Pasumarthi KB, Field LJ (2002) Cardiomyocyte cell cycle regulation. Circ Res 90: 1044–1054. - PubMed
-
- Walsh S, Ponten A, Fleischmann BK, Jovinge S (2010) Cardiomyocyte cell cycle control and growth estimation in vivo–an analysis based on cardiomyocyte nuclei. Cardiovasc Res 86: 365–373. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
