

Hsp70 oligomerization is mediated by an interaction between the interdomain linker and the substrate-binding domain
- PMID: 23840795
- PMCID: PMC3696110
- DOI: 10.1371/journal.pone.0067961
Hsp70 oligomerization is mediated by an interaction between the interdomain linker and the substrate-binding domain
Abstract
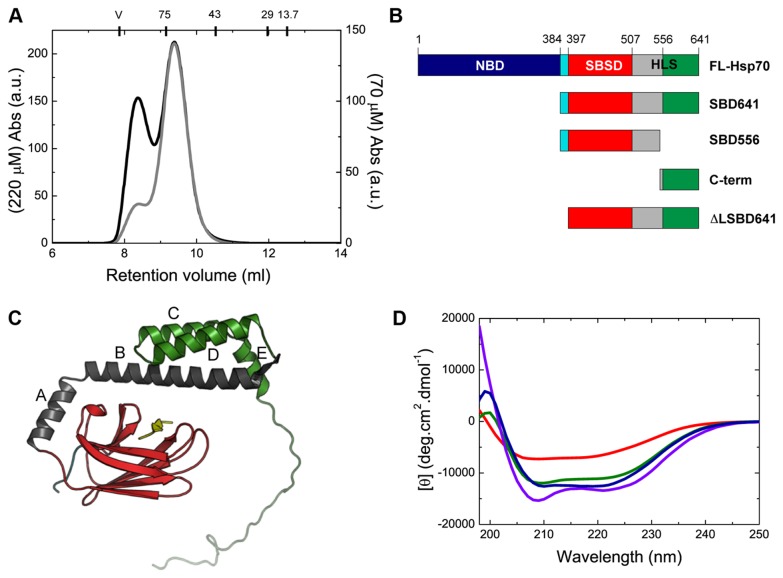
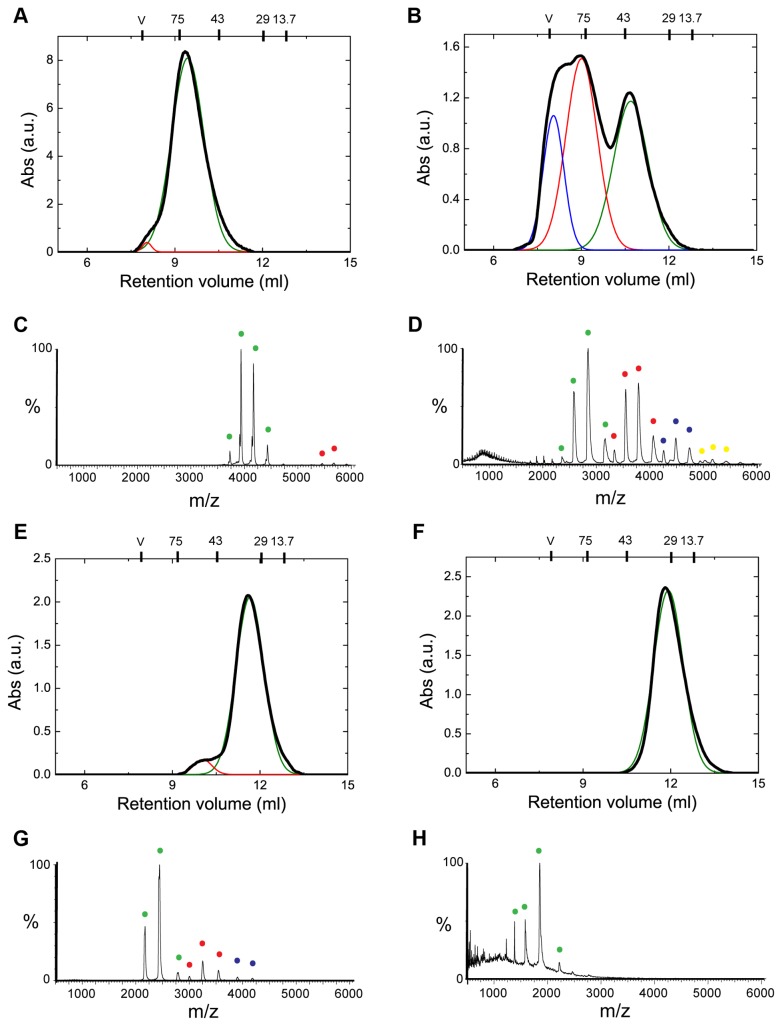
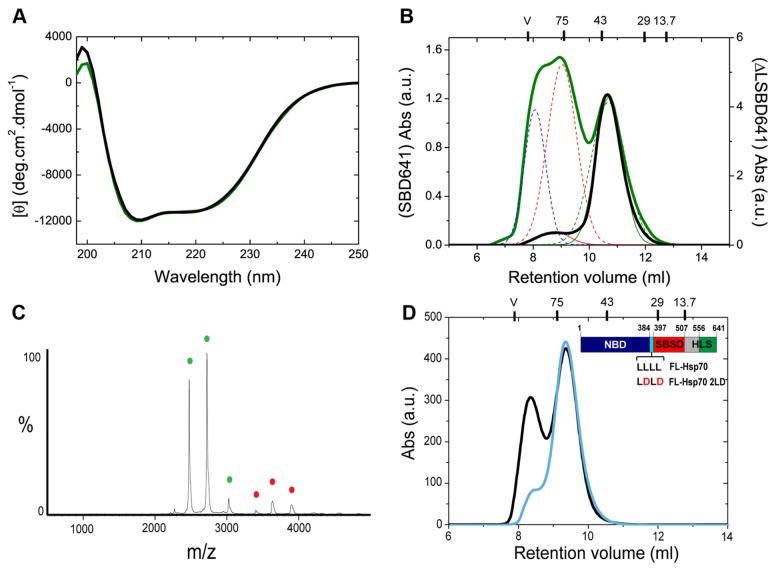
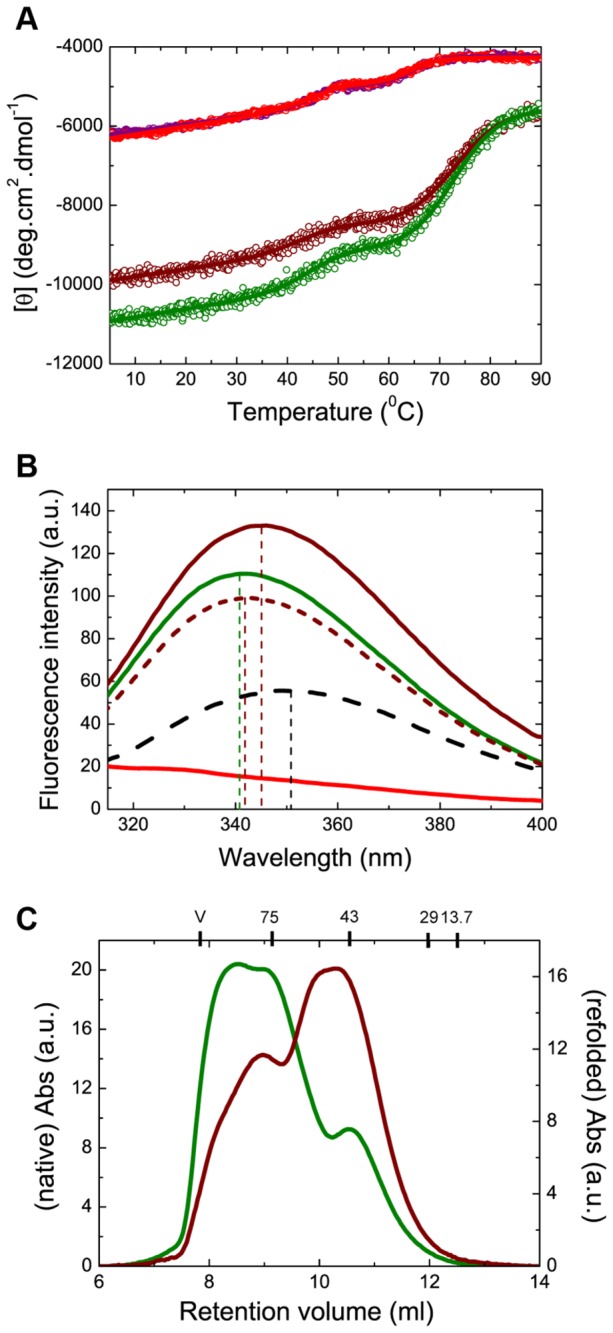
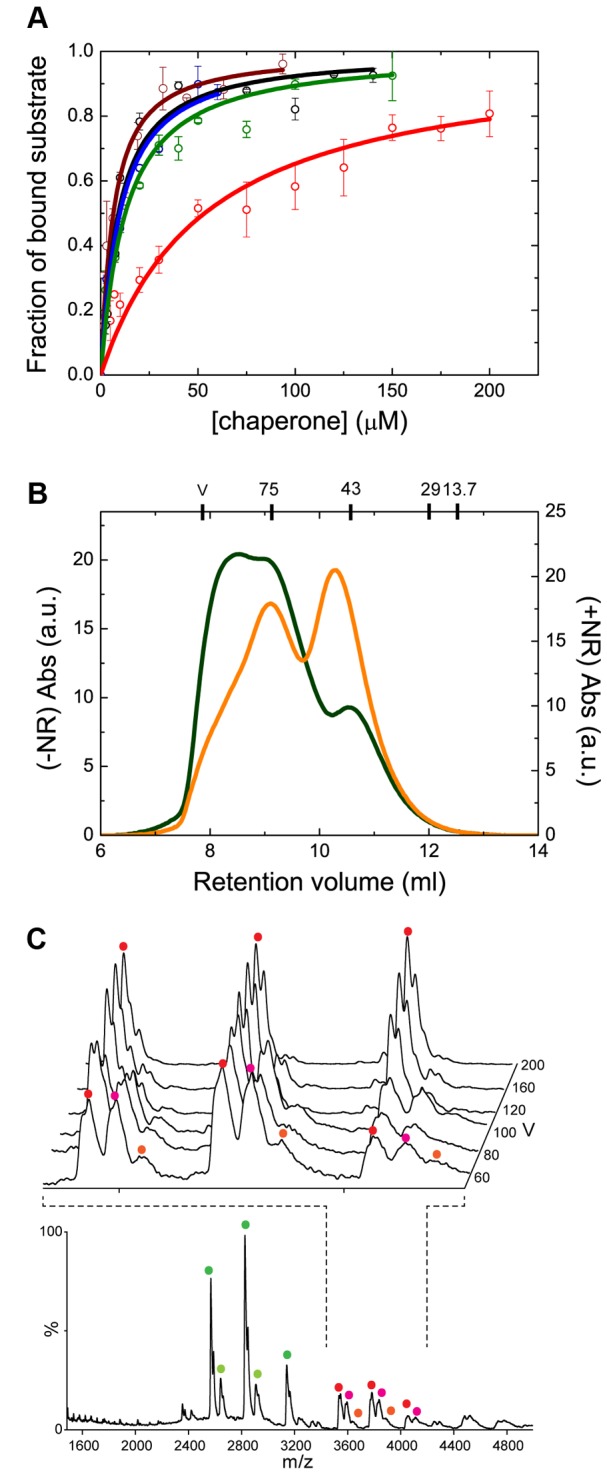
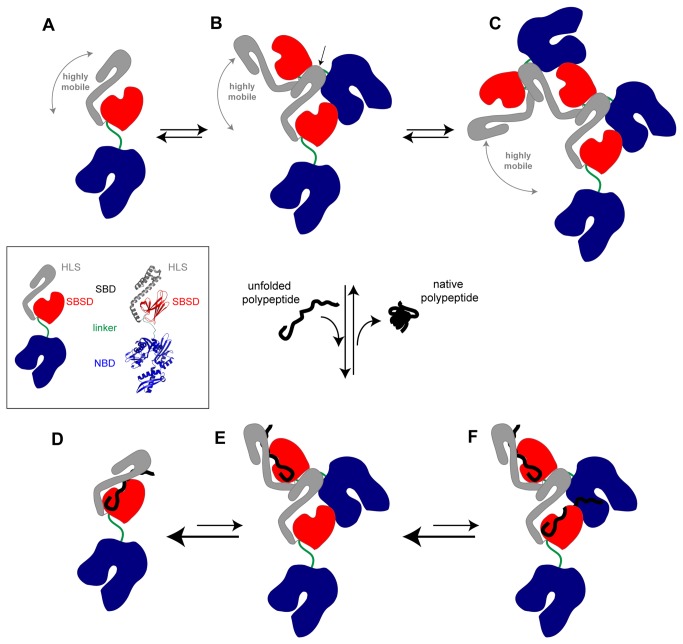
Oligomerization in the heat shock protein (Hsp) 70 family has been extensively documented both in vitro and in vivo, although the mechanism, the identity of the specific protein regions involved and the physiological relevance of this process are still unclear. We have studied the oligomeric properties of a series of human Hsp70 variants by means of nanoelectrospray ionization mass spectrometry, optical spectroscopy and quantitative size exclusion chromatography. Our results show that Hsp70 oligomerization takes place through a specific interaction between the interdomain linker of one molecule and the substrate-binding domain of a different molecule, generating dimers and higher-order oligomers. We have found that substrate binding shifts the oligomerization equilibrium towards the accumulation of functional monomeric protein, probably by sequestering the helical lid sub-domain needed to stabilize the chaperone: substrate complex. Taken together, these findings suggest a possible role of chaperone oligomerization as a mechanism for regulating the availability of the active monomeric form of the chaperone and for the control of substrate binding and release.
Conflict of interest statement
Figures
References
-
- Evans CG, Chang L, Gestwicki JE (2010) Heat shock protein 70 (hsp70) as an emerging drug target. J Med Chem 53: 4585-4602. doi:10.1021/jm100054f. PubMed: 20334364. - DOI - PMC - PubMed
-
- Gupta RS, Singh B (1994) Phylogenetic analysis of 70 kD heat shock protein sequences suggests a chimeric origin for the eukaryotic cell nucleus. Curr Biol 4: 1104-1114. doi:10.1016/S0960-9822(00)00249-9. PubMed: 7704574. - DOI - PubMed
-
- Mayer MP, Schröder H, Rüdiger S, Paal K, Laufen T et al. (2000) Multistep mechanism of substrate binding determines chaperone activity of Hsp70. Nat Struct Biol 7: 586-593. - PubMed
-
- Vogel M, Mayer MP, Bukau B (2006) Allosteric regulation of Hsp70 chaperones involves a conserved interdomain linker. J Biol Chem 281: 38705-38711. doi:10.1074/jbc.M609020200. PubMed: 17052976. - DOI - PubMed
-
- Liu Q, Hendrickson WA (2007) Insights into Hsp70 chaperone activity from a crystal structure of the yeast Hsp110 Sse1. Cell 131: 106-120. doi:10.1016/j.cell.2007.08.039. PubMed: 17923091. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
