

A longitudinal study of Caenorhabditis elegans larvae reveals a novel locomotion switch, regulated by G(αs) signaling
- PMID: 23840929
- PMCID: PMC3699835
- DOI: 10.7554/eLife.00782
A longitudinal study of Caenorhabditis elegans larvae reveals a novel locomotion switch, regulated by G(αs) signaling
Abstract
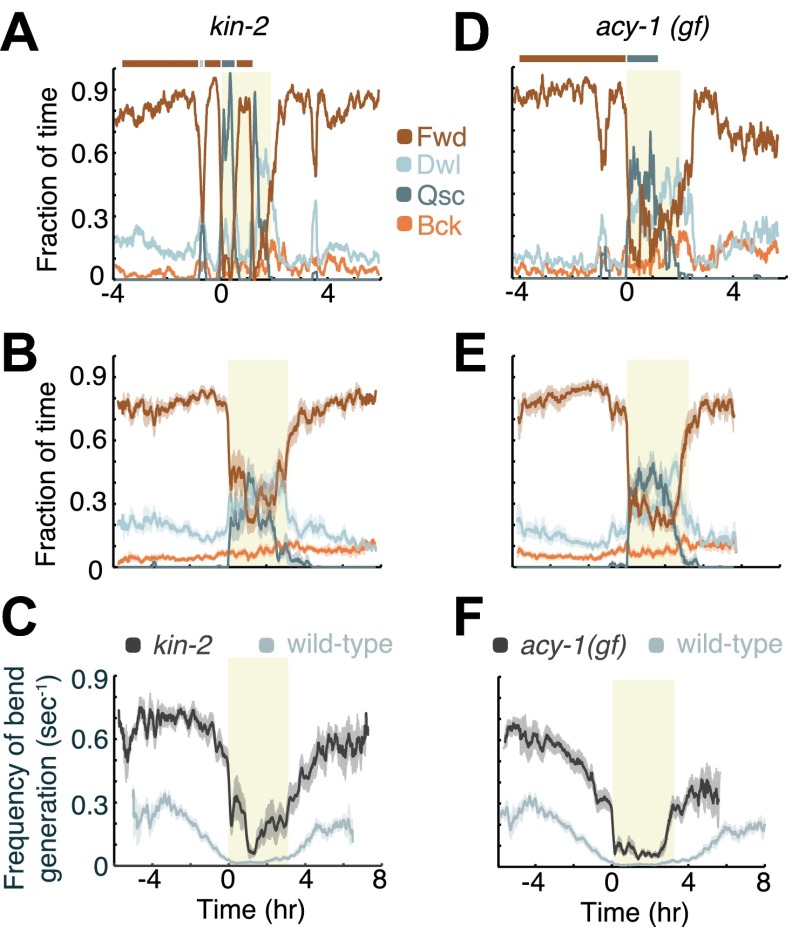
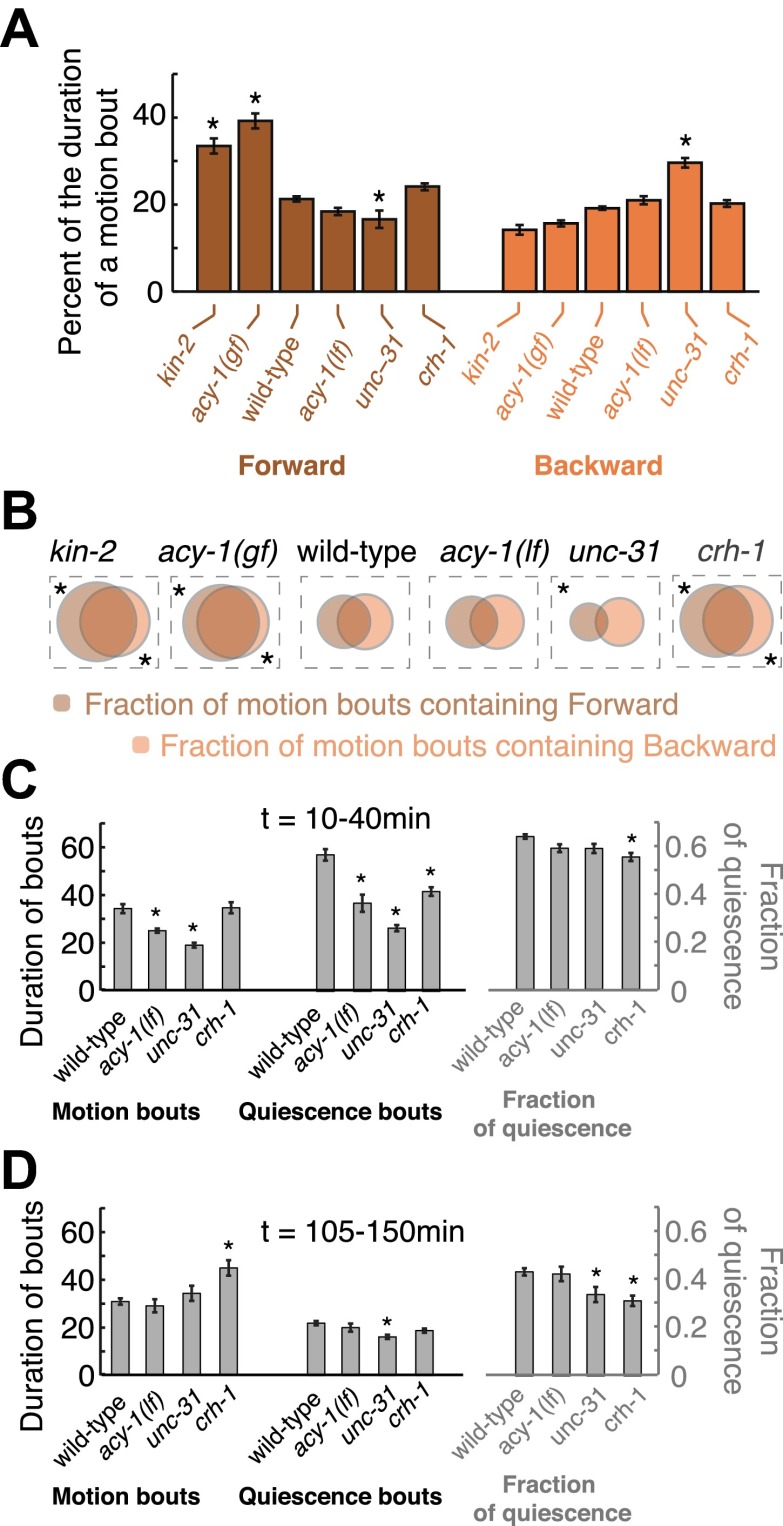
Despite their simplicity, longitudinal studies of invertebrate models are rare. We thus sought to characterize behavioral trends of Caenorhabditis elegans, from the mid fourth larval stage through the mid young adult stage. We found that, outside of lethargus, animals exhibited abrupt switching between two distinct behavioral states: active wakefulness and quiet wakefulness. The durations of epochs of active wakefulness exhibited non-Poisson statistics. Increased Gαs signaling stabilized the active wakefulness state before, during and after lethargus. In contrast, decreased Gαs signaling, decreased neuropeptide release, or decreased CREB activity destabilized active wakefulness outside of, but not during, lethargus. Taken together, our findings support a model in which protein kinase A (PKA) stabilizes active wakefulness, at least in part through two of its downstream targets: neuropeptide release and CREB. However, during lethargus, when active wakefulness is strongly suppressed, the native role of PKA signaling in modulating locomotion and quiescence may be minor. DOI:http://dx.doi.org/10.7554/eLife.00782.001.
Keywords: C. elegans; CREB; PKA; longitudinal study; modulation; quiet wakefulness; unc-31/CAPS.
Conflict of interest statement
The authors declare that no competing interests exist.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
