

Targeting of the tumor necrosis factor receptor superfamily for cancer immunotherapy
- PMID: 23840967
- PMCID: PMC3693168
- DOI: 10.1155/2013/371854
Targeting of the tumor necrosis factor receptor superfamily for cancer immunotherapy
Abstract
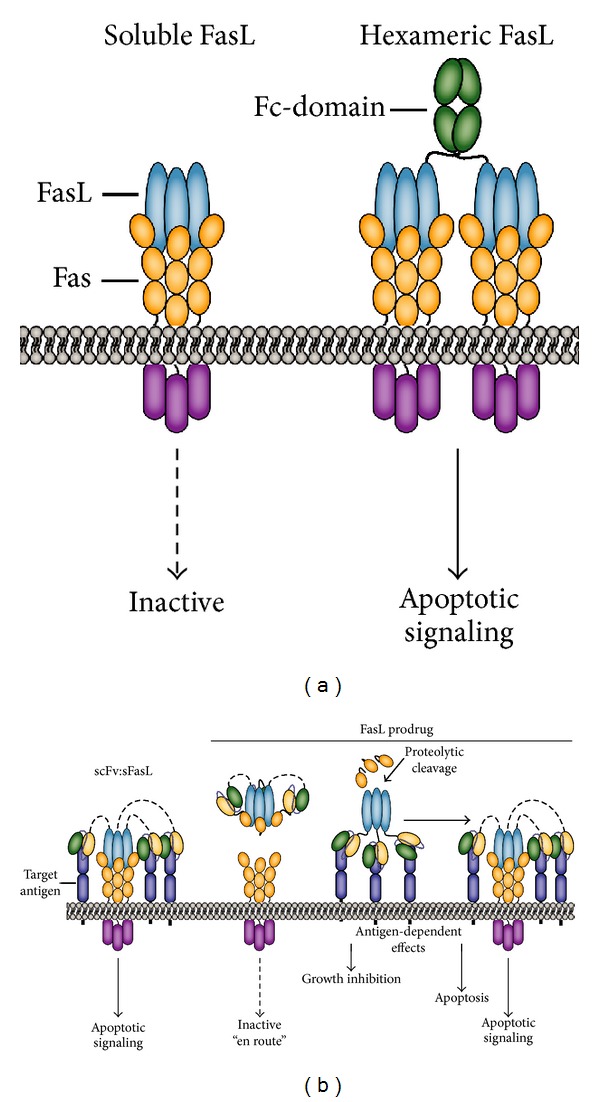
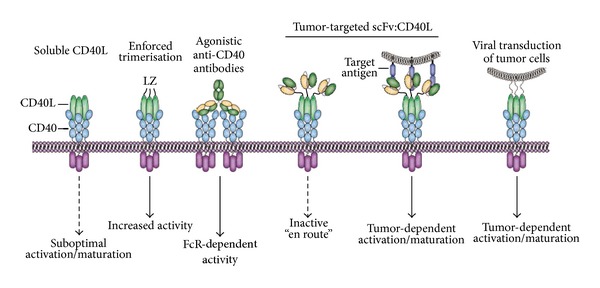
The tumor necrosis factor (TNF) ligand and cognate TNF receptor superfamilies constitute an important regulatory axis that is pivotal for immune homeostasis and correct execution of immune responses. TNF ligands and receptors are involved in diverse biological processes ranging from the selective induction of cell death in potentially dangerous and superfluous cells to providing costimulatory signals that help mount an effective immune response. This diverse and important regulatory role in immunity has sparked great interest in the development of TNFL/TNFR-targeted cancer immunotherapeutics. In this review, I will discuss the biology of the most prominent proapoptotic and co-stimulatory TNF ligands and review their current status in cancer immunotherapy.
Figures







References
-
- Banner DW, D’Arcy A, Janes W, et al. Crystal structure of the soluble human 55 kd TNF receptor-human TNFβ complex: implications for TNF receptor activation. Cell. 1993;73(3):431–445. - PubMed
-
- Naismith JH, Sprang SR. Modularity in the TNF-receptor family. Trends in Biochemical Sciences. 1998;23(2):74–79. - PubMed
-
- Peter ME, Krammer PH. The CD95(APO-1/Fas) DISC and beyond. Cell Death and Differentiation. 2003;10(1):26–35. - PubMed
-
- Kataoka T. The caspase-8 modulator c-FLIP. Critical Reviews in Immunology. 2005;25(1):31–58. - PubMed
-
- Boatright KM, Renatus M, Scott FL, et al. A unified model for apical caspase activation. Molecular Cell. 2003;11(2):529–541. - PubMed
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
