

Progress in targeting cell envelope biogenesis in Mycobacterium tuberculosis
- PMID: 23841633
- PMCID: PMC3867987
- DOI: 10.2217/fmb.13.52
Progress in targeting cell envelope biogenesis in Mycobacterium tuberculosis
Abstract
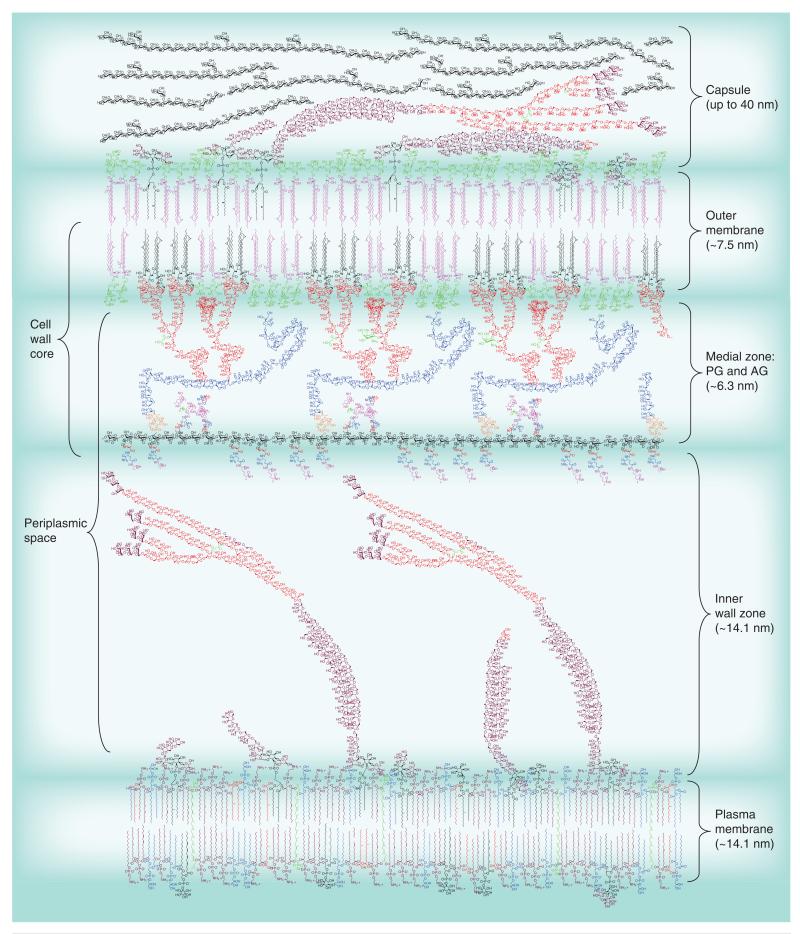
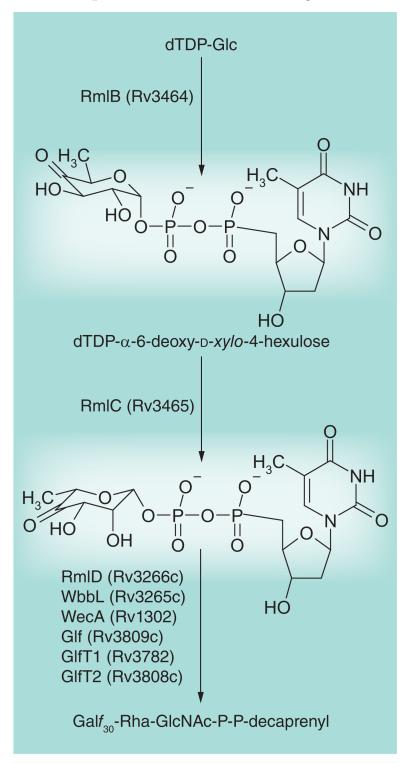
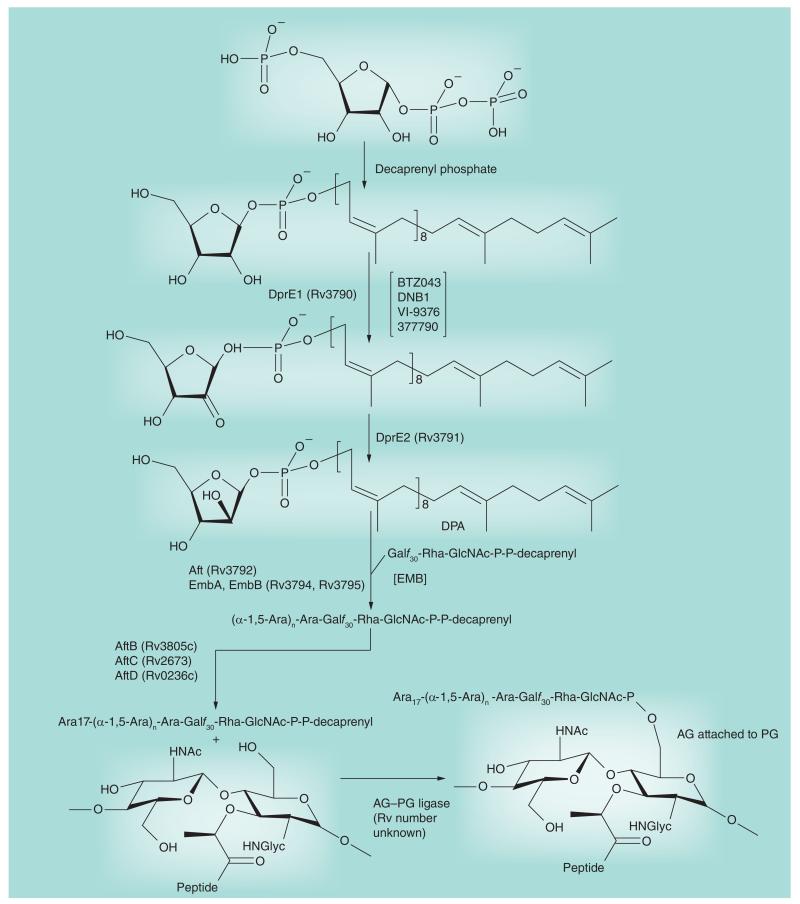
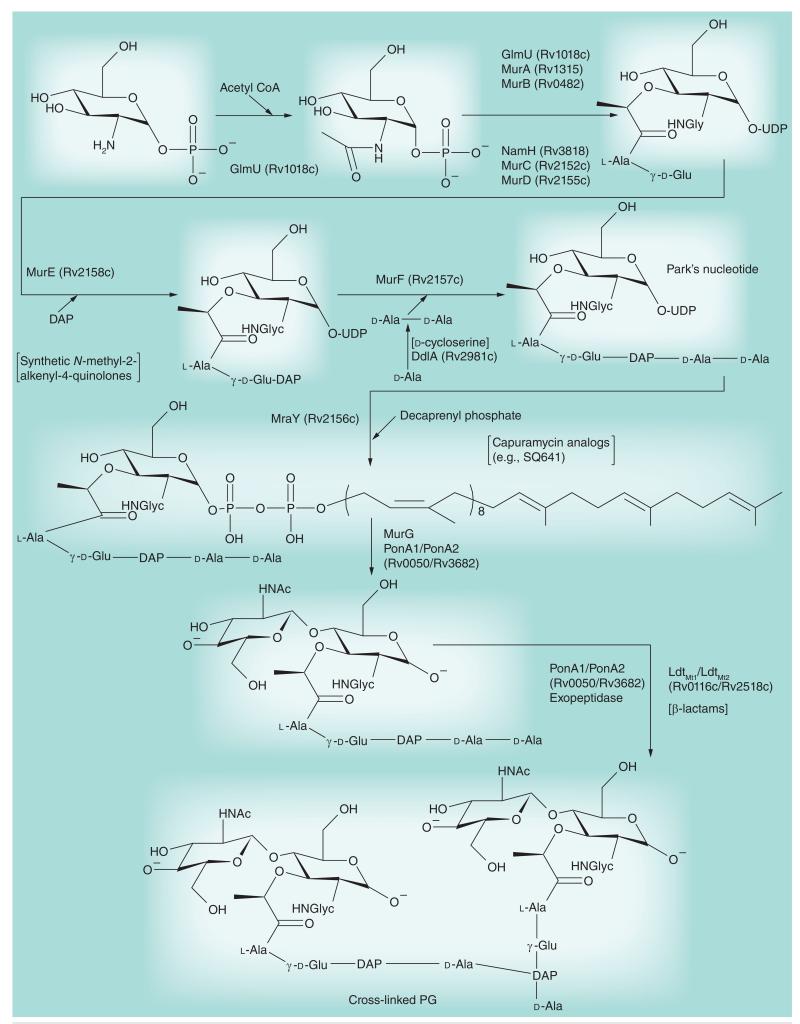
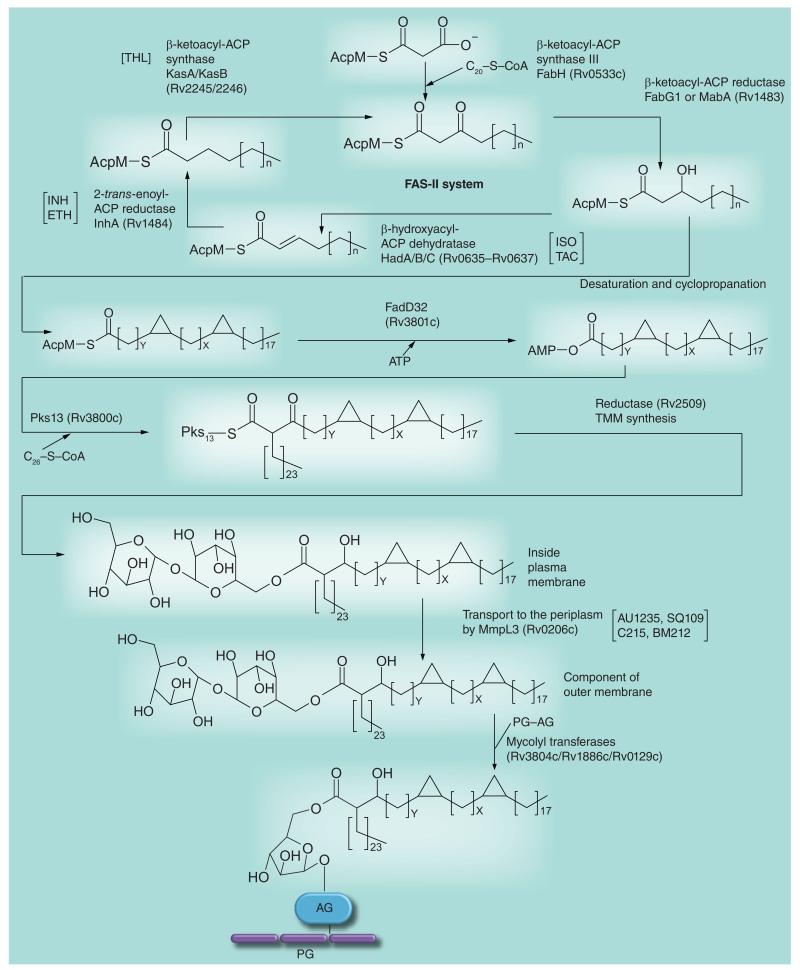
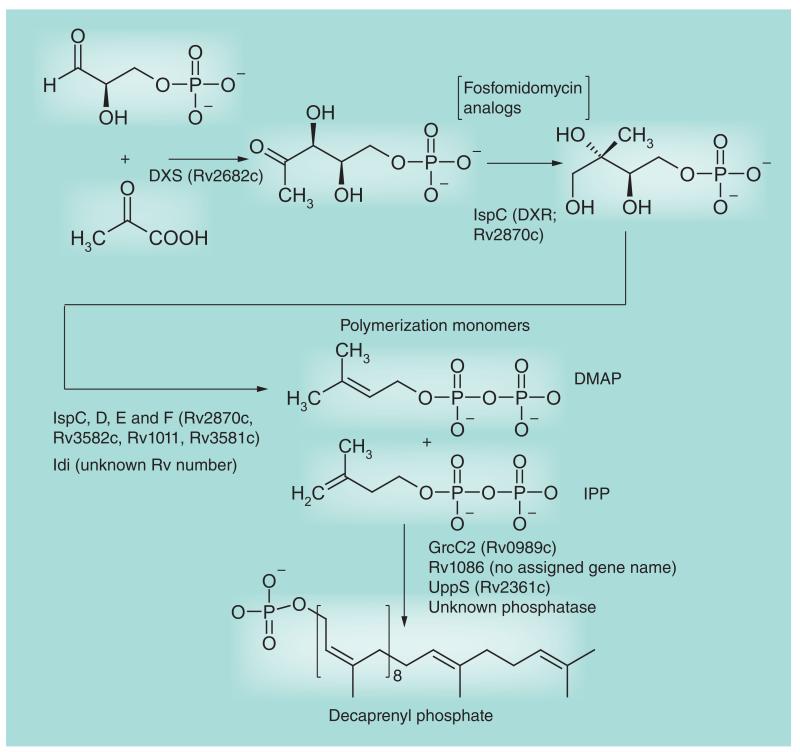
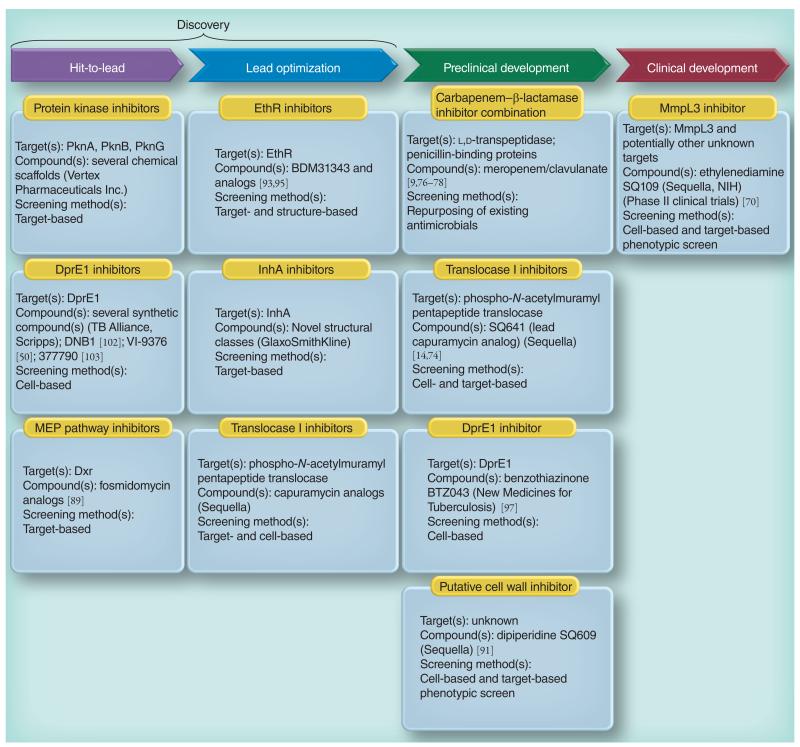
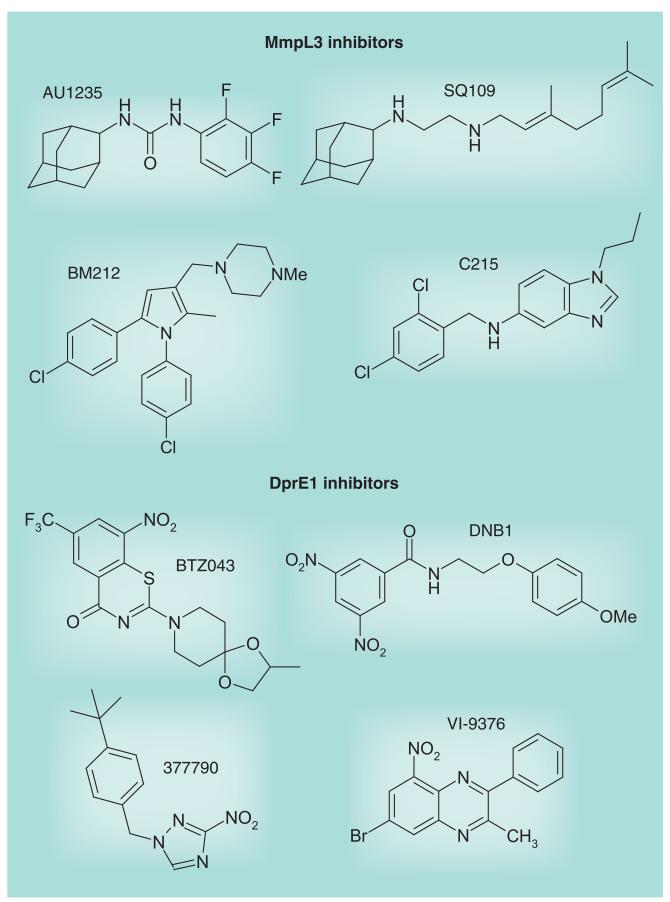
Most of the newly discovered compounds showing promise for the treatment of TB, notably multidrug-resistant TB, inhibit aspects of Mycobacterium tuberculosis cell envelope metabolism. This review reflects on the evolution of the knowledge that many of the front-line and emerging products inhibit aspects of cell envelope metabolism and in the process are bactericidal not only against actively replicating M. tuberculosis, but contrary to earlier impressions, are effective against latent forms of the disease. While mycolic acid and arabinogalactan synthesis are still primary targets of existing and new drugs, peptidoglycan synthesis, transport mechanisms and the synthesis of the decaprenyl-phosphate carrier lipid all show considerable promise as targets for new products, older drugs and new combinations. The advantages of whole cell- versus target-based screening in the perpetual search for new targets and products to counter multidrug-resistant TB are discussed.
Figures
References
-
- Center for Disease Control and Prevention: emergence of Mycobacterium tuberculosis with extensive resistance to second-line drugs worldwide, 2000-2004. MMWR Morb. Mortal. Wkly Rep. 2006;55(11):301–305. - PubMed
-
- Bhamidi S, Shi L, Chatterjee D, Belisle JT, Crick DC, McNeil MR. A bioanalytical method to determine the cell wall composition of Mycobacterium tuberculosis grown in vivo. Anal. Biochem. 2012;421(1):240–249. - PubMed
Websites
-
- WHO Tuberculosis. www.who.int/mediacentre/factsheets/fs104/en.
-
- PubChem BioAssay Inhibitors of mycobacterial glucosamine-1-phosphate acetyl transferase (GlmU) – bioassay summary. http://pubchem.ncbi.nlm.nih.gov/assay/assay.cgi?aid=1376.
-
- Working Group on New TB Drugs Stop TB Partnership. www.newtbdrugs.org.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical