

RrmA regulates the stability of specific transcripts in response to both nitrogen source and oxidative stress
- PMID: 23841692
- PMCID: PMC4282371
- DOI: 10.1111/mmi.12324
RrmA regulates the stability of specific transcripts in response to both nitrogen source and oxidative stress
Abstract
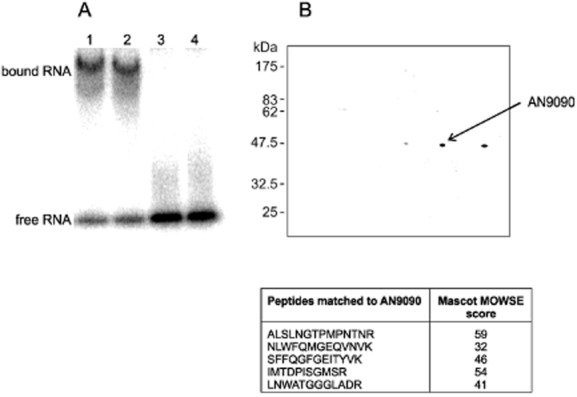
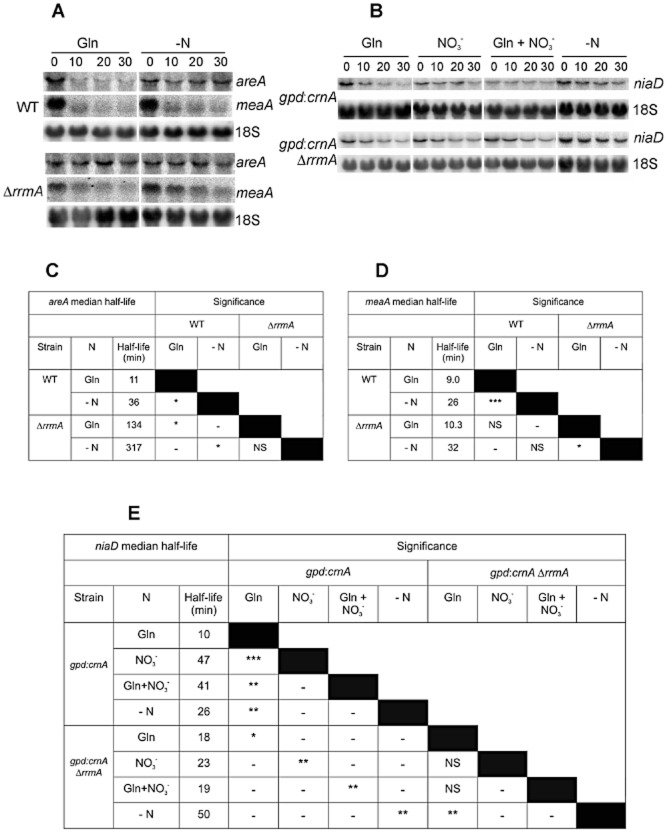
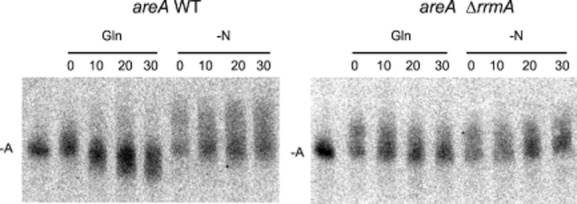
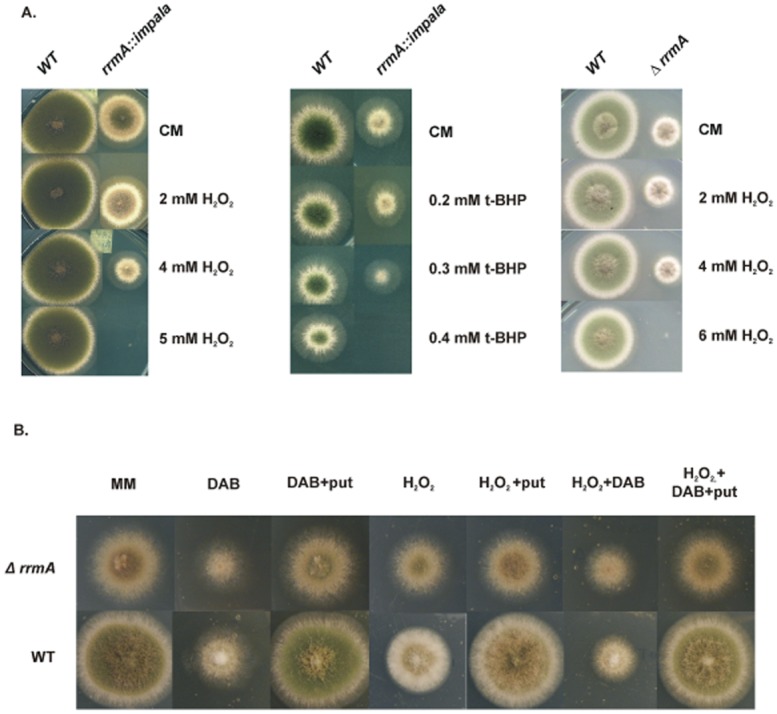
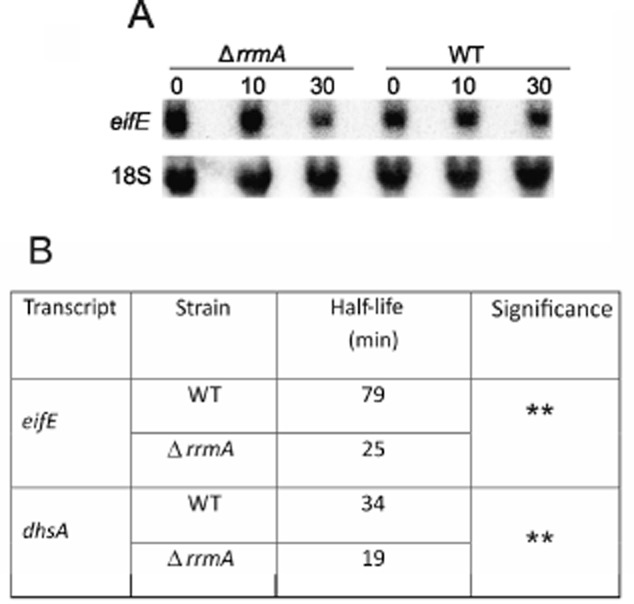
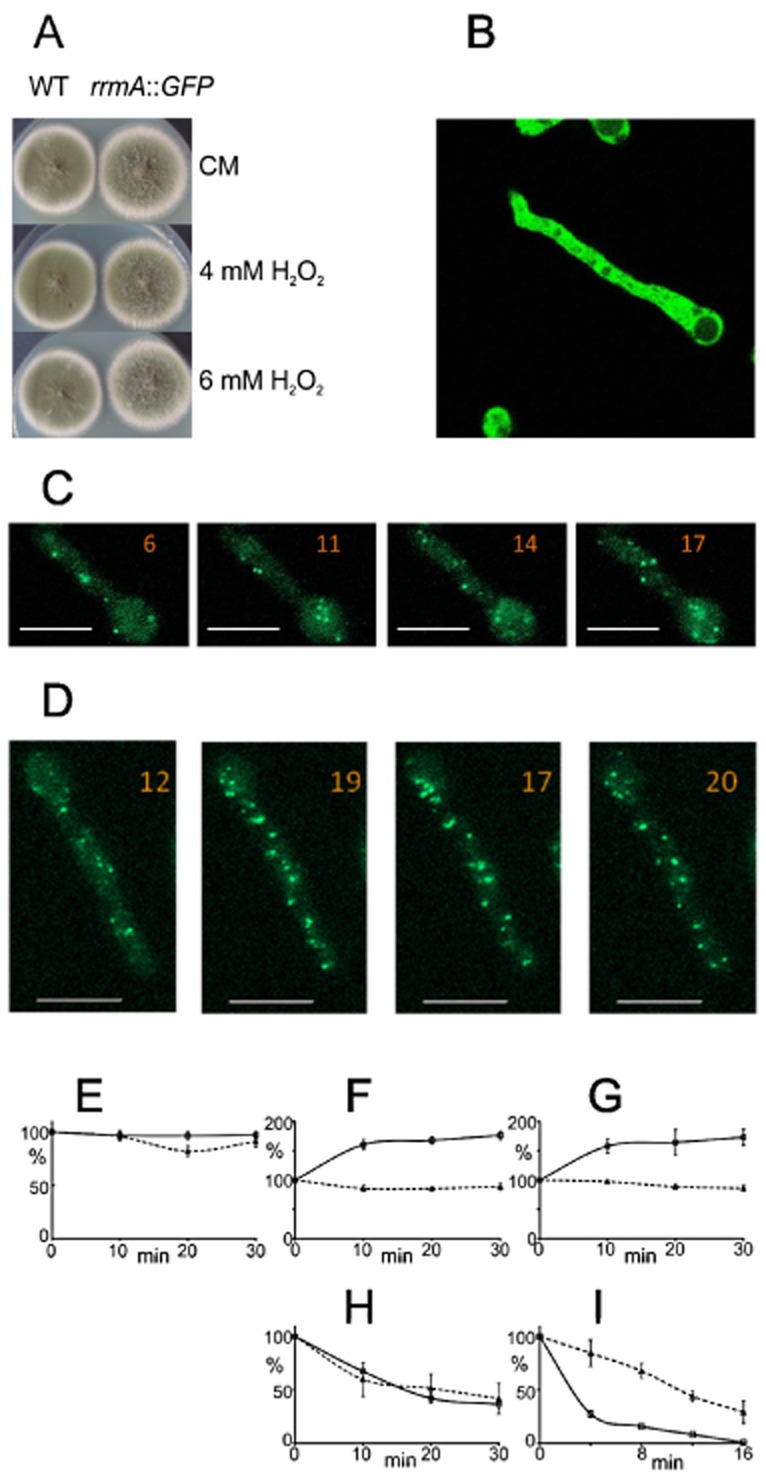
Differential regulation of transcript stability is an effective means by which an organism can modulate gene expression. A well-characterized example is glutamine signalled degradation of specific transcripts in Aspergillus nidulans. In the case of areA, which encodes a wide-domain transcription factor mediating nitrogen metabolite repression, the signal is mediated through a highly conserved region of the 3' UTR. Utilizing this RNA sequence we isolated RrmA, an RNA recognition motif protein. Disruption of the respective gene led to loss of both glutamine signalled transcript degradation as well as nitrate signalled stabilization of niaD mRNA. However, nitrogen starvation was shown to act independently of RrmA in stabilizing certain transcripts. RrmA was also implicated in the regulation of arginine catabolism gene expression and the oxidative stress responses at the level of mRNA stability. ΔrrmA mutants are hypersensitive to oxidative stress. This phenotype correlates with destabilization of eifE and dhsA mRNA. eifE encodes eIF5A, a translation factor within which a conserved lysine is post-translationally modified to hypusine, a process requiring DhsA. Intriguingly, for specific transcripts RrmA mediates both stabilization and destabilization and the specificity of the signals transduced is transcript dependent, suggesting it acts in consort with other factors which differ between transcripts.
© 2013 The Authors. Molecular Microbiology published by John Wiley & Sons Ltd.
Figures
References
-
- Asano Y, Hagiwara D, Yamashino T. Mizuno T. Characterization of the bZip-type transcription factor NapA with reference to oxidative stress response in Aspergillus nidulans. Biosci Biotechnol Biochem. 2007;71:1800–1803. - PubMed
-
- Baker CS, Morozov I, Suzuki K, Romeo T. Babitzke P. CsrA regulates glycogen biosynthesis by preventing translation of glgC in Escherichia coli. Mol Microbiol. 2002;44:1599–1610. - PubMed
-
- Beelman CA. Parker R. Differential effects of translational inhibition in cis and in trans on the decay of the unstable yeast MFA2 mRNA. J Biol Chem. 1994;269:9687–9692. - PubMed
-
- Bevilacqua A, Ceriani MC, Capaccioli S. Nicolin A. Post-transcriptional regulation of gene expression by degradation of messenger RNAs. J Cell Physiol. 2003;195:356–372. - PubMed
-
- Borsuk P, Przykorska A, Blachnio K, Koper M, Pawlowicz JM, Pekala M. Weglenski P. L-arginine influences the structure and function of arginase mRNA in Aspergillus nidulans. Biol Chem. 2007;388:135–144. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
