

Structure and function of the Salmonella Typhi chimaeric A(2)B(5) typhoid toxin
- PMID: 23842500
- PMCID: PMC4144355
- DOI: 10.1038/nature12377
Structure and function of the Salmonella Typhi chimaeric A(2)B(5) typhoid toxin
Abstract
Salmonella enterica serovar Typhi (S. Typhi) differs from most other salmonellae in that it causes a life-threatening systemic infection known as typhoid fever. The molecular bases for its unique clinical presentation are unknown. Here we find that the systemic administration of typhoid toxin, a unique virulence factor of S. Typhi, reproduces many of the acute symptoms of typhoid fever in an animal model. We identify specific carbohydrate moieties on specific surface glycoproteins that serve as receptors for typhoid toxin, which explains its broad cell target specificity. We present the atomic structure of typhoid toxin, which shows an unprecedented A2B5 organization with two covalently linked A subunits non-covalently associated to a pentameric B subunit. The structure provides insight into the toxin's receptor-binding specificity and delivery mechanisms and reveals how the activities of two powerful toxins have been co-opted into a single, unique toxin that can induce many of the symptoms characteristic of typhoid fever. These findings may lead to the development of potentially life-saving therapeutics against typhoid fever.
Conflict of interest statement
The authors declare no financial competing interests. Readers are welcome to comment on the on line version of the manuscript.
Figures
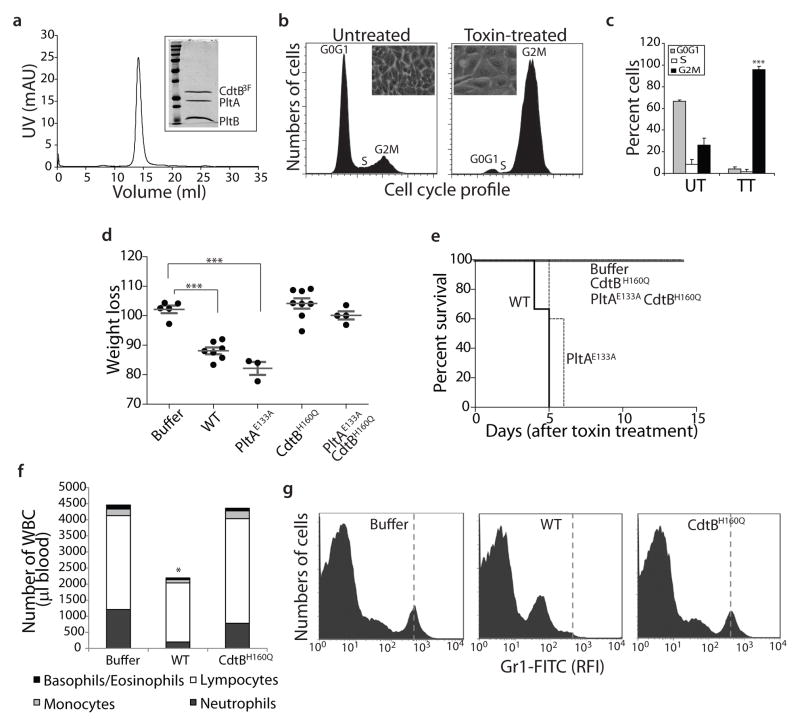
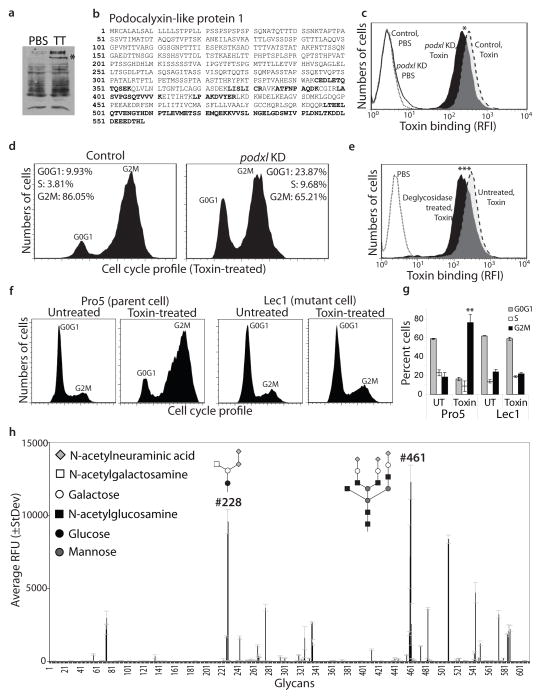
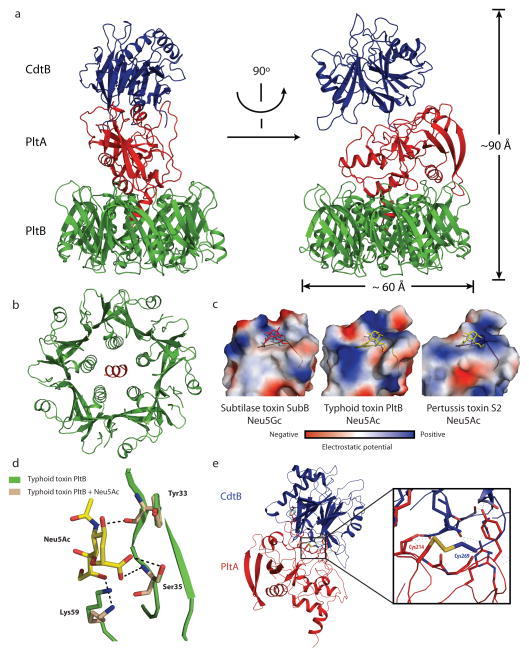
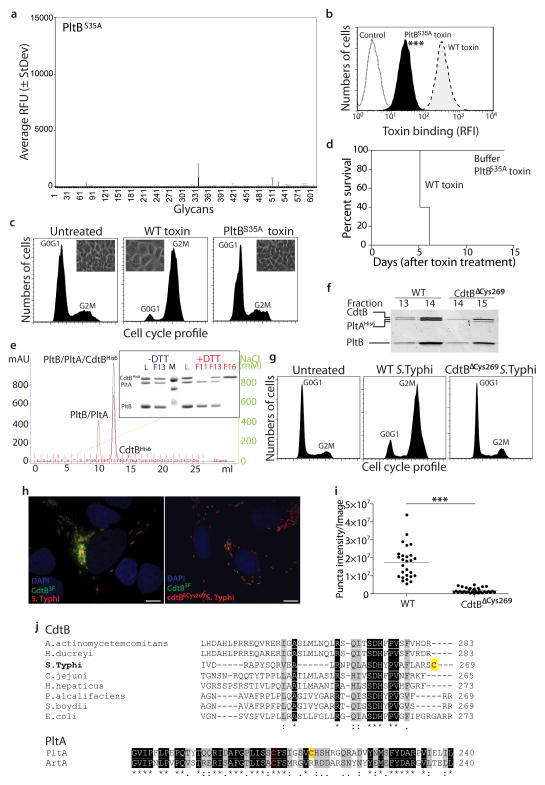
Comment in
-
Bacteriology: toxins in tandem.Nature. 2013 Jul 18;499(7458):293. doi: 10.1038/nature12409. Epub 2013 Jul 10. Nature. 2013. PMID: 23842489 No abstract available.
References
-
- Parry C, Hien TT, Dougan G, White N, Farrar J. Typhoid fever. N Engl J Med. 2002;347:1770–1782. - PubMed
-
- Parkhill J, et al. Complete genome sequence of a multiple drug resistant Salmonella enterica serovar Typhi CT18. Nature. 2001;413:848–852. - PubMed
-
- Butler T. Treatment of typhoid fever in the 21st century: promises and shortcomings. Clin Microbiol Infect. 2011;17:959–963. - PubMed
-
- Sabbagh S, Forest C, Lepage C, Leclerc J, Daigle F. So similar, yet so different: uncovering distinctive features in the genomes of Salmonella enterica serovars Typhimurium and Typhi. FEMS Microbiol Lett. 2010;305:1–13. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
