

The length-tension curve in muscle depends on lattice spacing
- PMID: 23843386
- PMCID: PMC3730583
- DOI: 10.1098/rspb.2013.0697
The length-tension curve in muscle depends on lattice spacing
Abstract
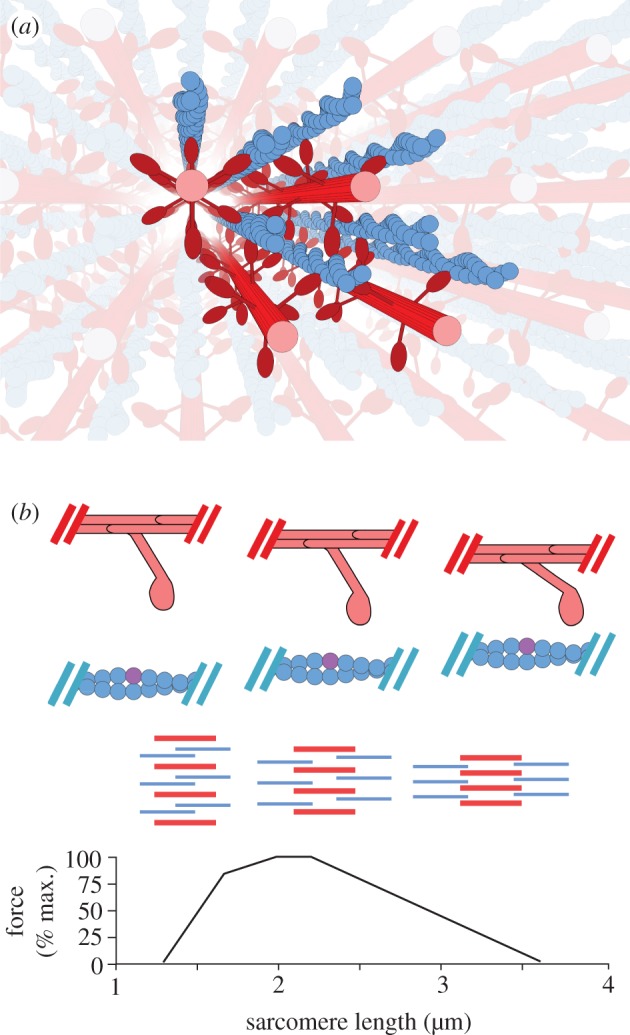
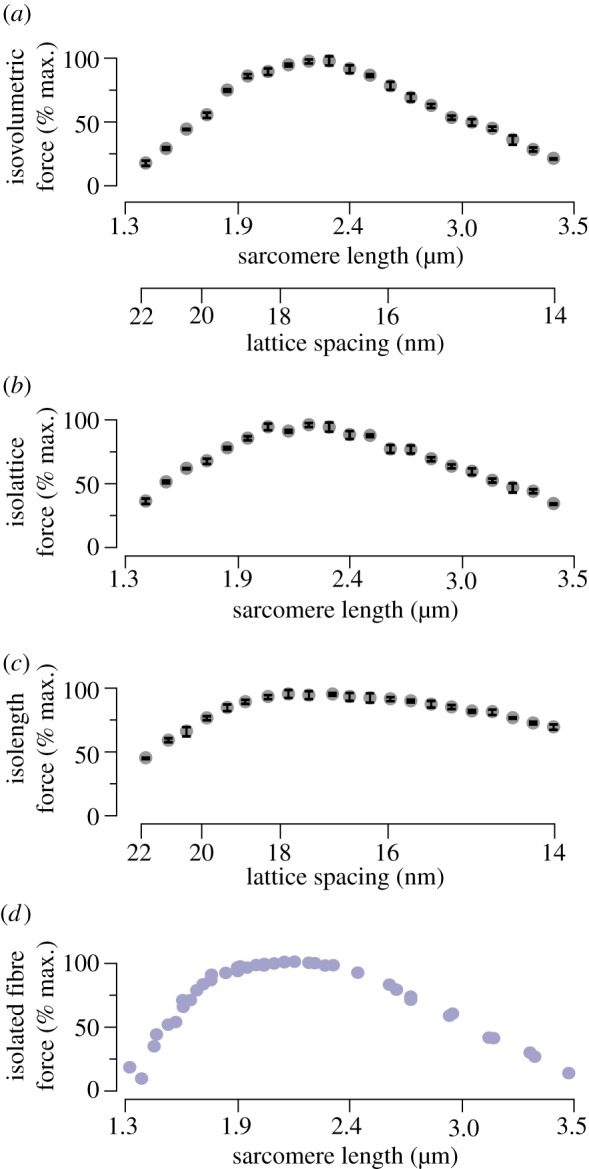
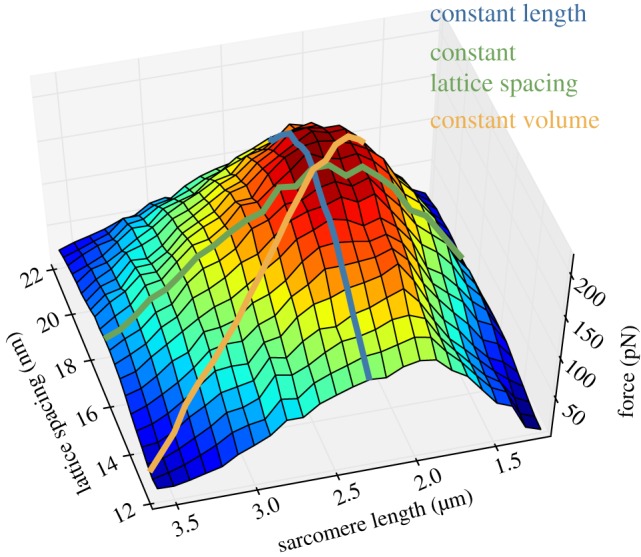
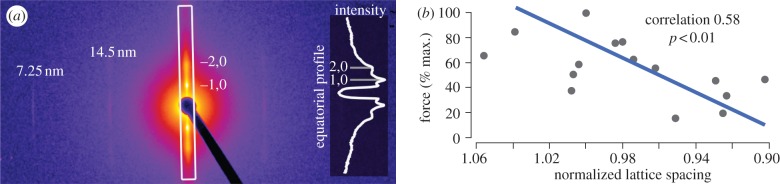
Classic interpretations of the striated muscle length-tension curve focus on how force varies with overlap of thin (actin) and thick (myosin) filaments. New models of sarcomere geometry and experiments with skinned synchronous insect flight muscle suggest that changes in the radial distance between the actin and myosin filaments, the filament lattice spacing, are responsible for between 20% and 50% of the change in force seen between sarcomere lengths of 1.4 and 3.4 µm. Thus, lattice spacing is a significant force regulator, increasing the slope of muscle's force-length dependence.
Keywords: X-ray diffraction; lattice spacing; length–tension curve; muscle contraction; spatially explicit.
Figures
References
-
- Cobb M. 2002. Timeline: exorcizing the animal spirits: Jan Swammerdam on nerve function. Nat. Rev. Neurosci. 3, 395–400 (doi:10.1038/nrn806) - DOI - PubMed
-
- Irving TC, Konhilas JP, Perry D, Fischetti R, de Tombe PP. 2000. Myofilament lattice spacing as a function of sarcomere length in isolated rat myocardium. Am. J. Physiol. Heart Circ. Physiol. 279, H2568–H2573 - PubMed
-
- Millman BM. 1998. The filament lattice of striated muscle. Physiol. Rev. 78, 359–391 - PubMed
-
- Rassier DE, MacIntosh BR, Herzog W. 1999. Length dependence of active force production in skeletal muscle. J. Appl. Physiol. 86, 1445–1457 - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources