

Dynamic modulation of spike timing-dependent calcium influx during corticostriatal upstates
- PMID: 23843436
- PMCID: PMC4042418
- DOI: 10.1152/jn.00232.2013
Dynamic modulation of spike timing-dependent calcium influx during corticostriatal upstates
Abstract
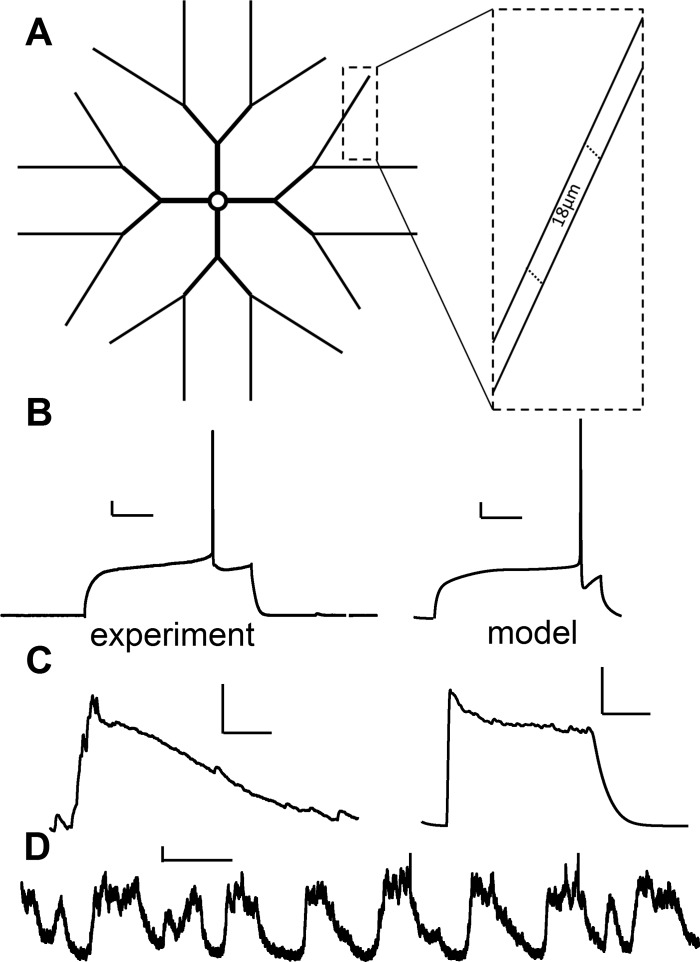
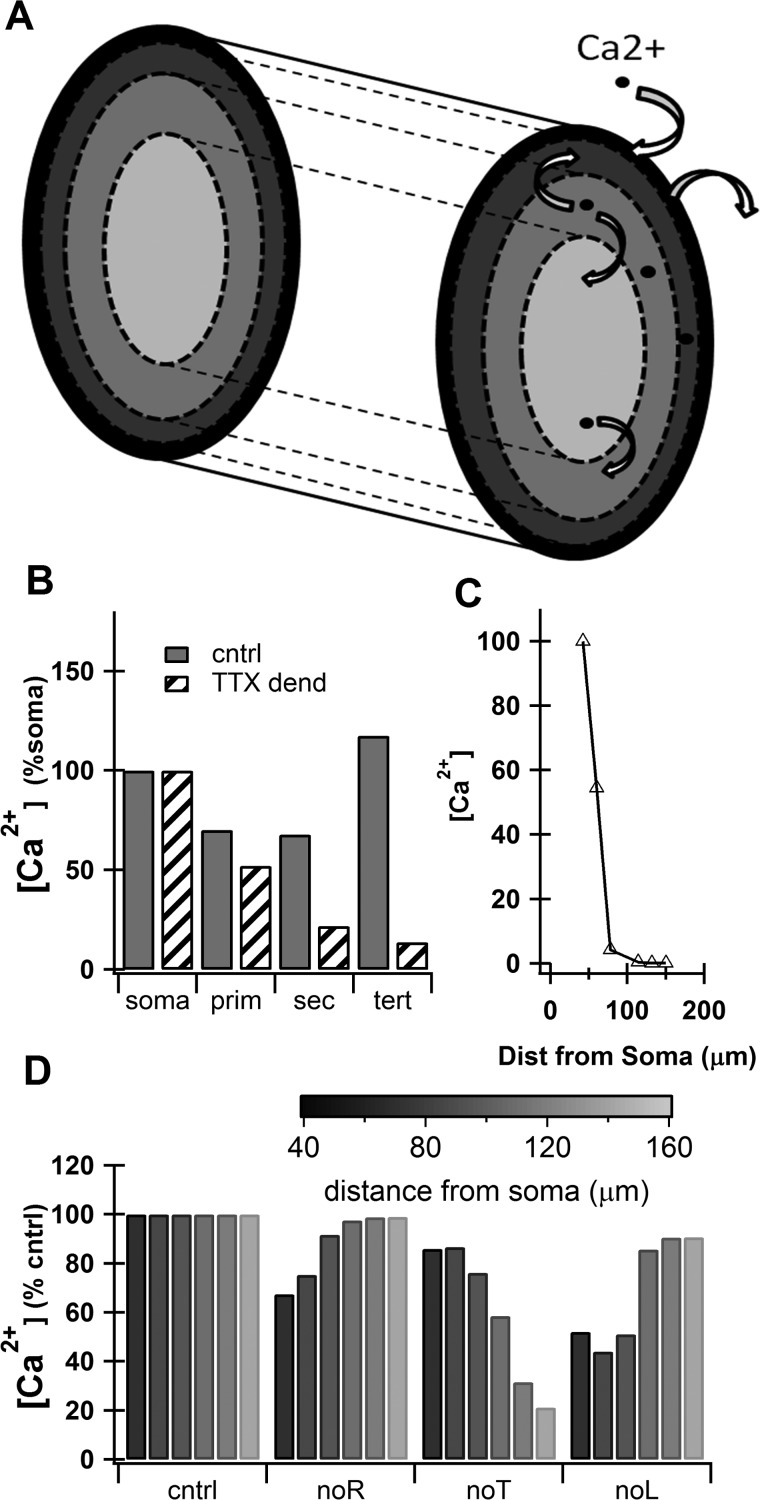
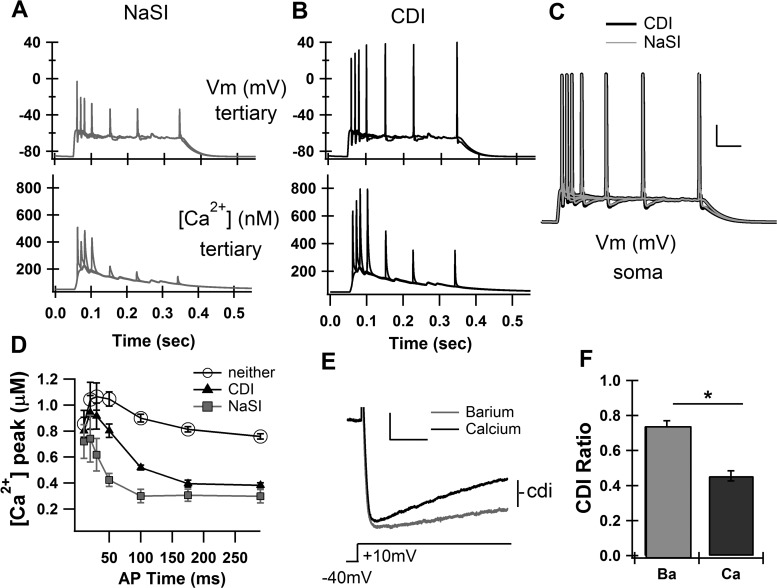
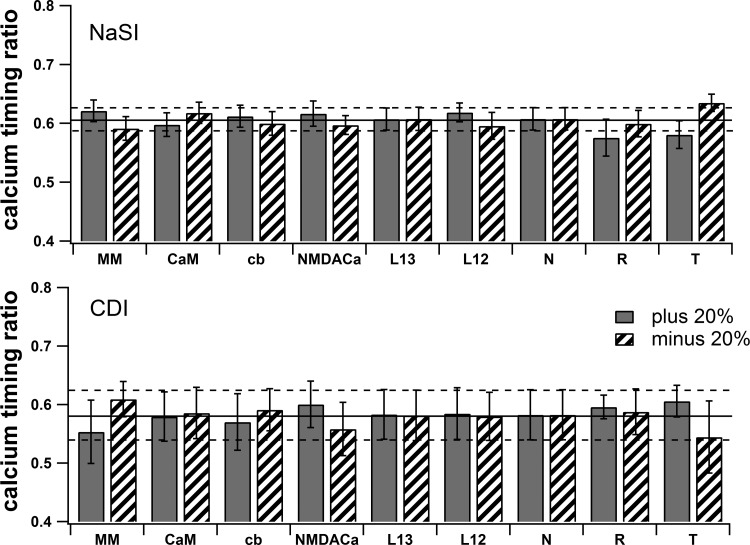
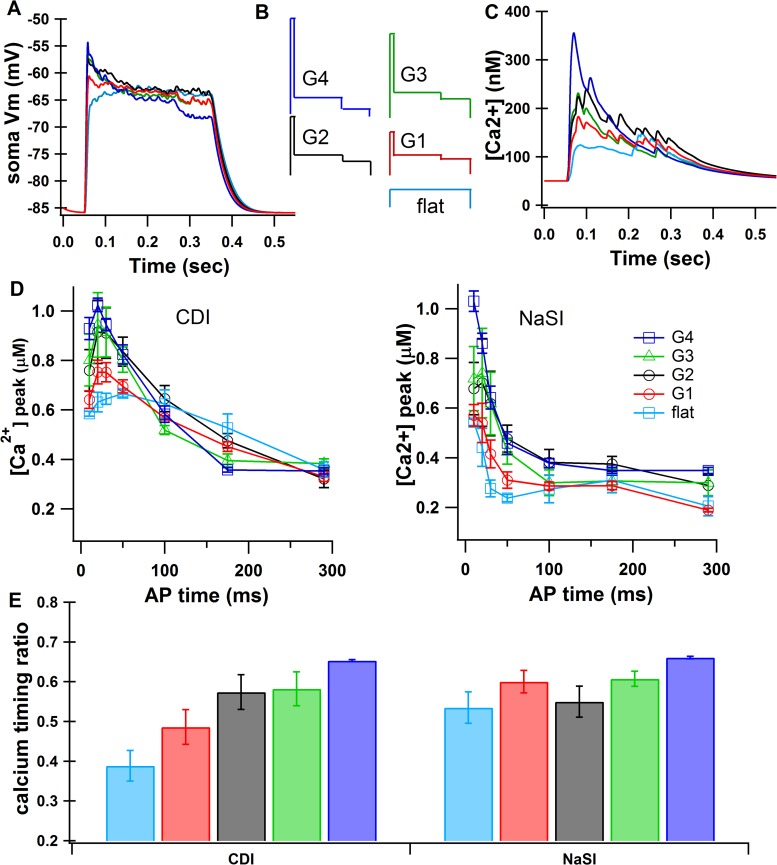
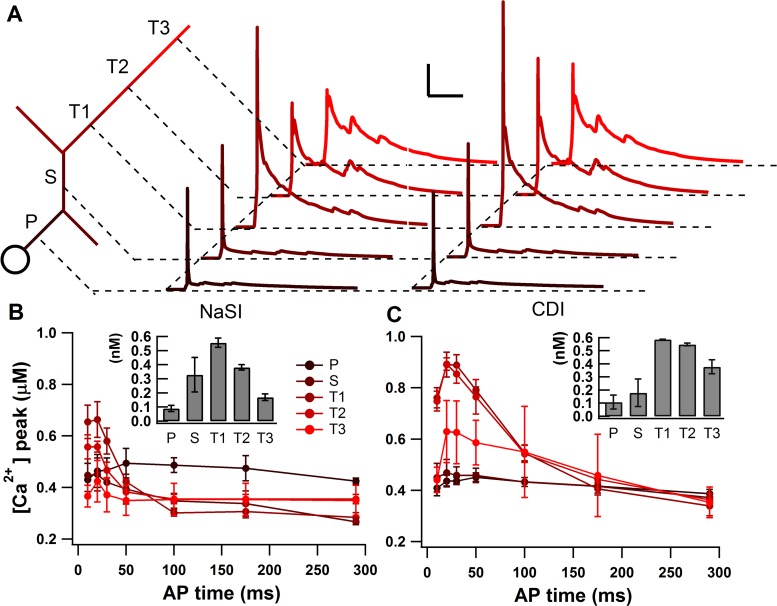
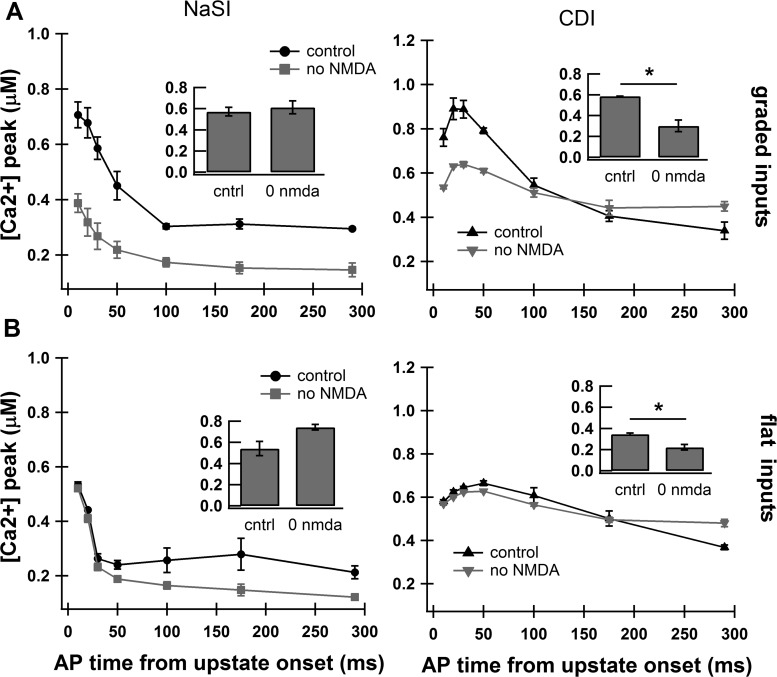
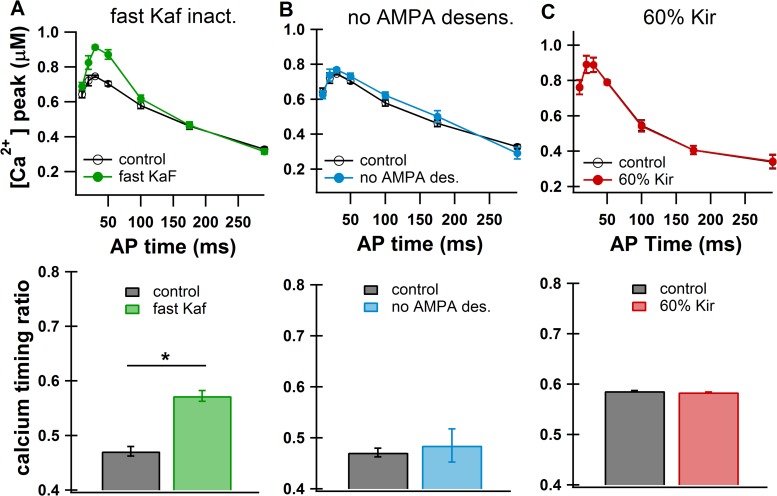
The striatum of the basal ganglia demonstrates distinctive upstate and downstate membrane potential oscillations during slow-wave sleep and under anesthetic. The upstates generate calcium transients in the dendrites, and the amplitude of these calcium transients depends strongly on the timing of the action potential (AP) within the upstate. Calcium is essential for synaptic plasticity in the striatum, and these large calcium transients during the upstates may control which synapses undergo plastic changes. To investigate the mechanisms that underlie the relationship between calcium and AP timing, we have developed a realistic biophysical model of a medium spiny neuron (MSN). We have implemented sophisticated calcium dynamics including calcium diffusion, buffering, and pump extrusion, which accurately replicate published data. Using this model, we found that either the slow inactivation of dendritic sodium channels (NaSI) or the calcium inactivation of voltage-gated calcium channels (CDI) can cause high calcium corresponding to early APs and lower calcium corresponding to later APs. We found that only CDI can account for the experimental observation that sensitivity to AP timing is dependent on NMDA receptors. Additional simulations demonstrated a mechanism by which MSNs can dynamically modulate their sensitivity to AP timing and show that sensitivity to specifically timed pre- and postsynaptic pairings (as in spike timing-dependent plasticity protocols) is altered by the timing of the pairing within the upstate. These findings have implications for synaptic plasticity in vivo during sleep when the upstate-downstate pattern is prominent in the striatum.
Keywords: calcium; computational model; medium spiny neuron; striatum; upstates.
Figures
Similar articles
-
Calcium dynamics predict direction of synaptic plasticity in striatal spiny projection neurons.Eur J Neurosci. 2017 Apr;45(8):1044-1056. doi: 10.1111/ejn.13287. Epub 2016 Jun 15. Eur J Neurosci. 2017. PMID: 27233469 Free PMC article.
-
Action potential timing determines dendritic calcium during striatal up-states.J Neurosci. 2004 Jan 28;24(4):877-85. doi: 10.1523/JNEUROSCI.4475-03.2004. J Neurosci. 2004. PMID: 14749432 Free PMC article.
-
A Ca(2+) threshold for induction of spike-timing-dependent depression in the mouse striatum.J Neurosci. 2011 Sep 7;31(36):13015-22. doi: 10.1523/JNEUROSCI.3206-11.2011. J Neurosci. 2011. PMID: 21900580 Free PMC article.
-
Spike timing, calcium signals and synaptic plasticity.Curr Opin Neurobiol. 2002 Jun;12(3):305-14. doi: 10.1016/s0959-4388(02)00325-2. Curr Opin Neurobiol. 2002. PMID: 12049938 Review.
-
Postsynaptic integration of glutamatergic and dopaminergic signals in the striatum.Prog Neurobiol. 1994 Oct;44(2):163-96. doi: 10.1016/0301-0082(94)90037-x. Prog Neurobiol. 1994. PMID: 7831476 Review.
Cited by
-
Synaptic cleft geometry modulates NMDAR opening probability by tuning neurotransmitter residence time.Biophys J. 2025 Apr 1;124(7):1058-1072. doi: 10.1016/j.bpj.2025.01.019. Epub 2025 Jan 28. Biophys J. 2025. PMID: 39876560 Free PMC article.
-
T2N as a new tool for robust electrophysiological modeling demonstrated for mature and adult-born dentate granule cells.Elife. 2017 Nov 22;6:e26517. doi: 10.7554/eLife.26517. Elife. 2017. PMID: 29165247 Free PMC article.
-
Calcium dynamics predict direction of synaptic plasticity in striatal spiny projection neurons.Eur J Neurosci. 2017 Apr;45(8):1044-1056. doi: 10.1111/ejn.13287. Epub 2016 Jun 15. Eur J Neurosci. 2017. PMID: 27233469 Free PMC article.
-
Functional Dissection of Basal Ganglia Inhibitory Inputs onto Substantia Nigra Dopaminergic Neurons.Cell Rep. 2020 Sep 15;32(11):108156. doi: 10.1016/j.celrep.2020.108156. Cell Rep. 2020. PMID: 32937133 Free PMC article.
-
A stochastic model of hippocampal synaptic plasticity with geometrical readout of enzyme dynamics.Elife. 2023 Aug 17;12:e80152. doi: 10.7554/eLife.80152. Elife. 2023. PMID: 37589251 Free PMC article.
References
-
- Allbritton NL, Meyer T, Stryer L. Range of messenger action of calcium ion and inositol 1,4,5-trisphosphate. Science 258: 1812–1815, 1992 - PubMed
-
- Berlanga ML, Price DL, Phung BS, Giuly R, Terada M, Yamada N, Cyr M, Caron MG, Laakso A, Martone ME, Ellisman MH. Multiscale imaging characterization of dopamine transporter knockout mice reveals regional alterations in spine density of medium spiny neurons. Brain Res 1390: 41–49, 2011 - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous