

Distinctive changes in plasma membrane phosphoinositides underlie differential regulation of TRPV1 in nociceptive neurons
- PMID: 23843517
- PMCID: PMC3724548
- DOI: 10.1523/JNEUROSCI.5637-12.2013
Distinctive changes in plasma membrane phosphoinositides underlie differential regulation of TRPV1 in nociceptive neurons
Abstract
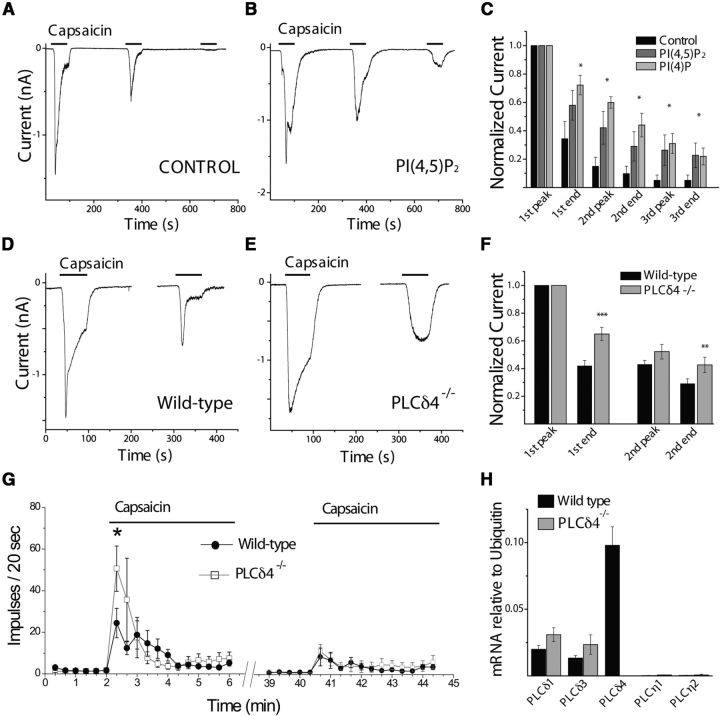
Transient Receptor Potential Vanilloid 1 (TRPV1) is a polymodal, Ca(2+)-permeable cation channel crucial to regulation of nociceptor responsiveness. Sensitization of TRPV1 by G-protein coupled receptor (GPCR) agonists to its endogenous activators, such as low pH and noxious heat, is a key factor in hyperalgesia during tissue injury as well as pathological pain syndromes. Conversely, chronic pharmacological activation of TRPV1 by capsaicin leads to calcium influx-induced adaptation of the channel. Paradoxically, both conditions entail activation of phospholipase C (PLC) enzymes, which hydrolyze phosphoinositides. We found that in sensory neurons PLCβ activation by bradykinin led to a moderate decrease in phosphatidylinositol-4,5-bisphosphate (PI(4,5)P2), but no sustained change in the levels of its precursor PI(4)P. Preventing this selective decrease in PI(4,5)P2 inhibited TRPV1 sensitization, while selectively decreasing PI(4,5)P2 independently of PLC potentiated the sensitizing effect of protein kinase C (PKC) on the channel, thereby inducing increased TRPV1 responsiveness. Maximal pharmacological TRPV1 stimulation led to a robust decrease of both PI(4,5)P2 and its precursor PI(4)P in sensory neurons. Attenuating the decrease of either lipid significantly reduced desensitization, and simultaneous reduction of PI(4,5)P2 and PI(4)P independently of PLC inhibited TRPV1. We found that, on the mRNA level, the dominant highly Ca(2+)-sensitive PLC isoform in dorsal root ganglia is PLCδ4. Capsaicin-induced desensitization of TRPV1 currents was significantly reduced, whereas capsaicin-induced nerve impulses in the skin-nerve preparation increased in mice lacking this isoform. We propose a comprehensive model in which differential changes in phosphoinositide levels mediated by distinct PLC isoforms result in opposing changes in TRPV1 activity.
Figures









Similar articles
-
Dual regulation of TRPV1 by phosphoinositides.J Neurosci. 2007 Jun 27;27(26):7070-80. doi: 10.1523/JNEUROSCI.1866-07.2007. J Neurosci. 2007. PMID: 17596456 Free PMC article.
-
Activation of TRPV1 channels inhibits mechanosensitive Piezo channel activity by depleting membrane phosphoinositides.Sci Signal. 2015 Feb 10;8(363):ra15. doi: 10.1126/scisignal.2005667. Sci Signal. 2015. PMID: 25670203 Free PMC article.
-
Diacylglycerol kinases regulate TRPV1 channel activity.J Biol Chem. 2020 Jun 12;295(24):8174-8185. doi: 10.1074/jbc.RA119.012505. Epub 2020 Apr 28. J Biol Chem. 2020. PMID: 32345612 Free PMC article.
-
Phosphoinositide regulation of TRPV1 revisited.Pflugers Arch. 2015 Sep;467(9):1851-69. doi: 10.1007/s00424-015-1695-3. Epub 2015 Mar 11. Pflugers Arch. 2015. PMID: 25754030 Free PMC article. Review.
-
Differential effects of TRPV channel block on polymodal activation of rat cutaneous nociceptors in vitro.Exp Brain Res. 2009 Jun;196(1):31-44. doi: 10.1007/s00221-009-1808-3. Epub 2009 Apr 30. Exp Brain Res. 2009. PMID: 19404626 Review.
Cited by
-
Regulation of the transient receptor potential channel TRPM3 by phosphoinositides.J Gen Physiol. 2015 Jul;146(1):51-63. doi: 10.1085/jgp.201411339. J Gen Physiol. 2015. PMID: 26123194 Free PMC article.
-
TRPV1: A Target for Rational Drug Design.Pharmaceuticals (Basel). 2016 Aug 23;9(3):52. doi: 10.3390/ph9030052. Pharmaceuticals (Basel). 2016. PMID: 27563913 Free PMC article. Review.
-
Ca2+ Regulation of TRP Ion Channels.Int J Mol Sci. 2018 Apr 23;19(4):1256. doi: 10.3390/ijms19041256. Int J Mol Sci. 2018. PMID: 29690581 Free PMC article. Review.
-
Hypersensitivity Induced by Intrathecal Bradykinin Administration Is Enhanced by N-oleoyldopamine (OLDA) and Prevented by TRPV1 Antagonist.Int J Mol Sci. 2021 Apr 2;22(7):3712. doi: 10.3390/ijms22073712. Int J Mol Sci. 2021. PMID: 33918267 Free PMC article.
-
Sensory TRP channels: the key transducers of nociception and pain.Prog Mol Biol Transl Sci. 2015;131:73-118. doi: 10.1016/bs.pmbts.2015.01.002. Epub 2015 Feb 12. Prog Mol Biol Transl Sci. 2015. PMID: 25744671 Free PMC article. Review.
References
-
- Bhave G, Hu HJ, Glauner KS, Zhu W, Wang H, Brasier DJ, Oxford GS, Gereau RW., 4th Protein kinase C phosphorylation sensitizes but does not activate the capsaicin receptor transient receptor potential vanilloid 1 (TRPV1) Proc Natl Acad Sci U S A. 2003;100:12480–12485. doi: 10.1073/pnas.2032100100. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous