

Predictive suppression of cortical excitability and its deficit in schizophrenia
- PMID: 23843536
- PMCID: PMC3724541
- DOI: 10.1523/JNEUROSCI.0010-13.2013
Predictive suppression of cortical excitability and its deficit in schizophrenia
Abstract
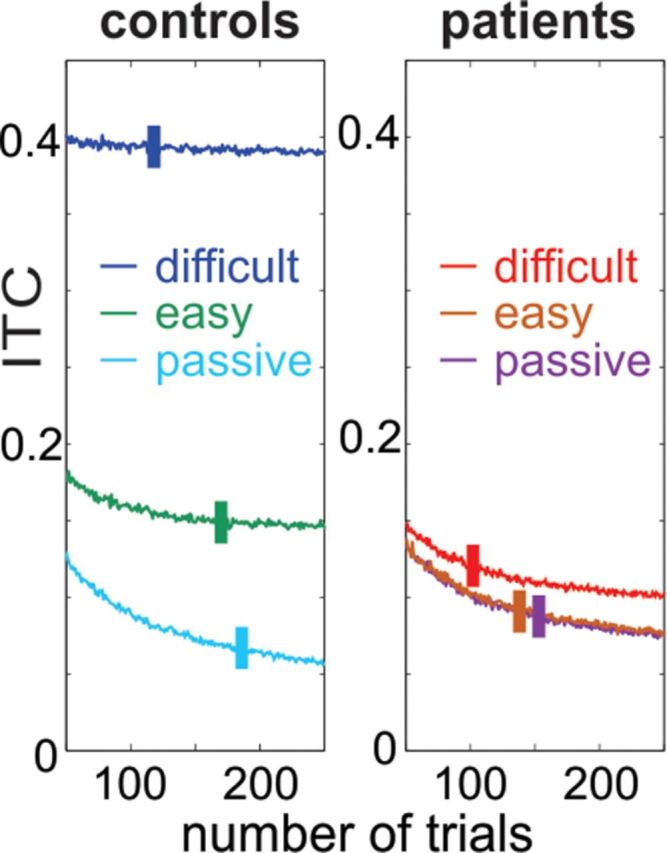
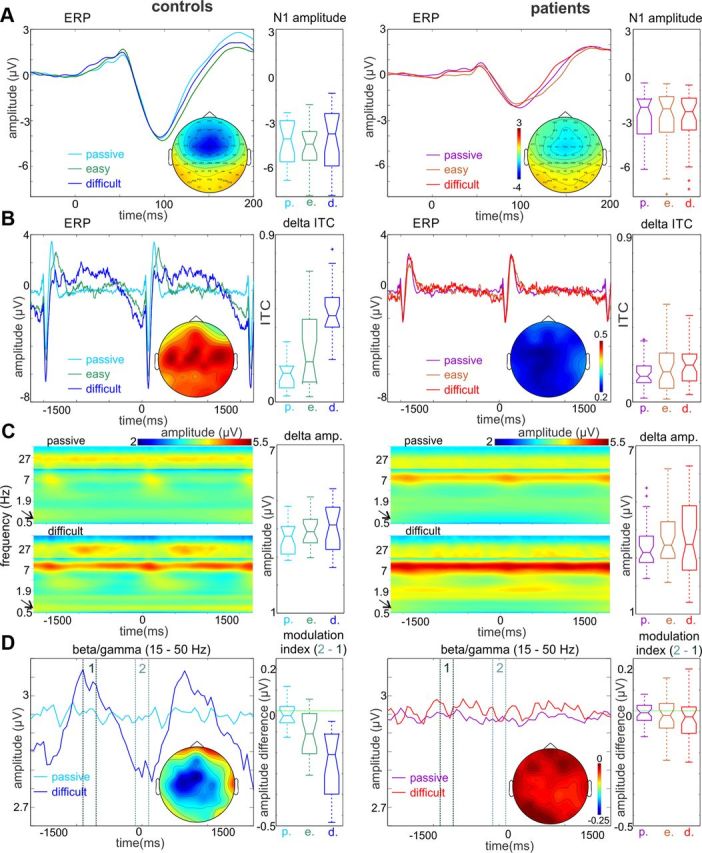
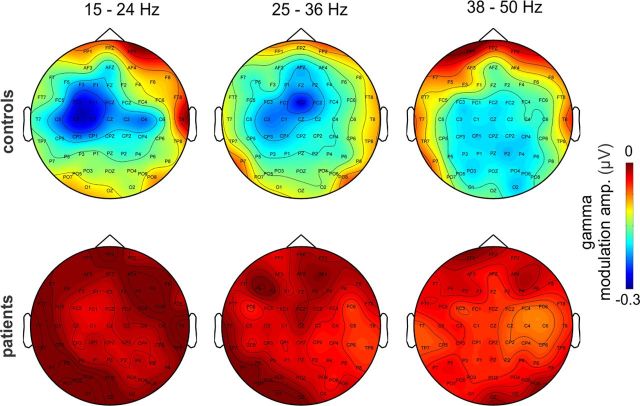
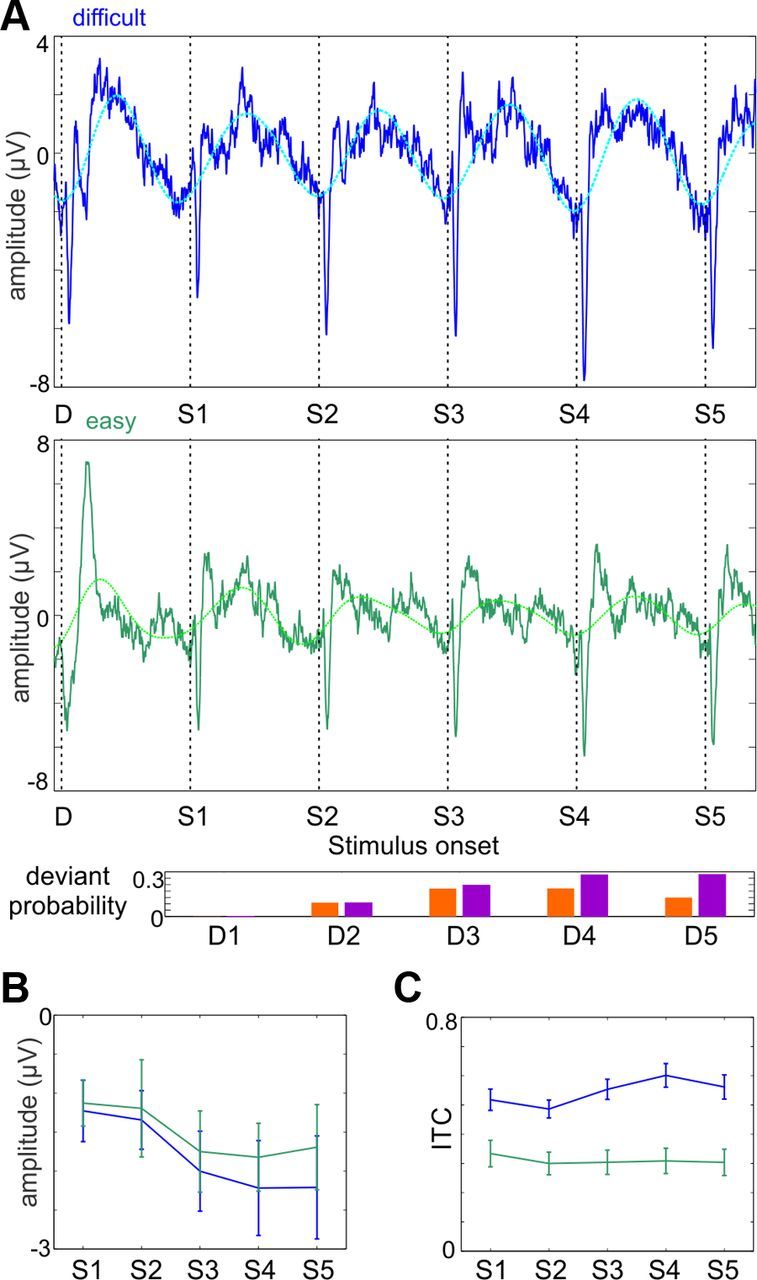
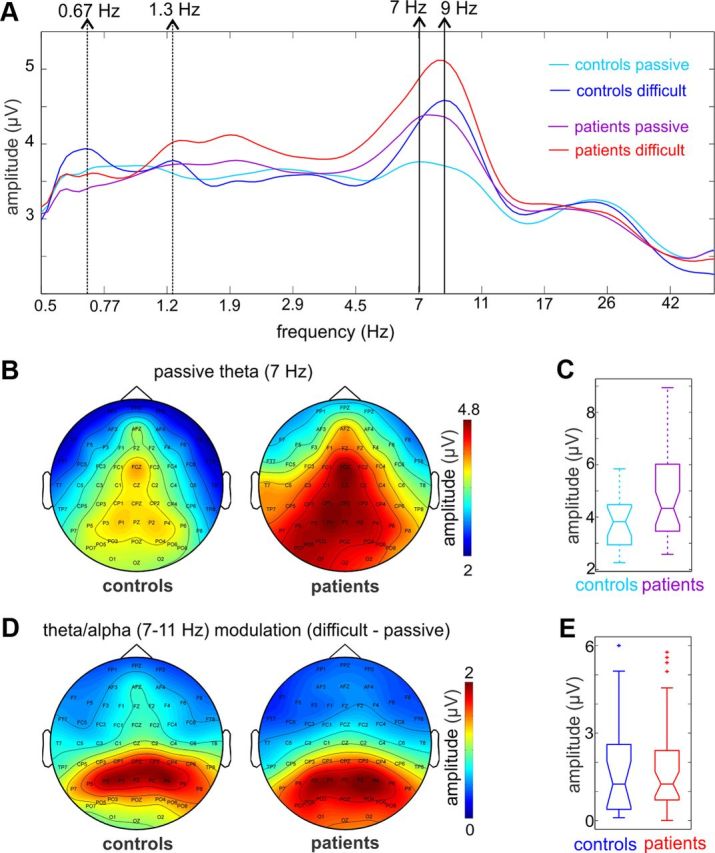
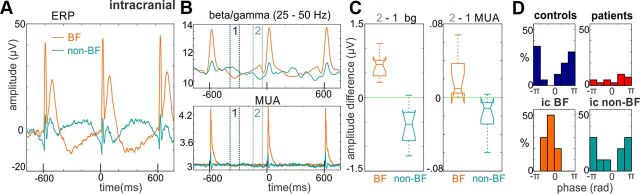
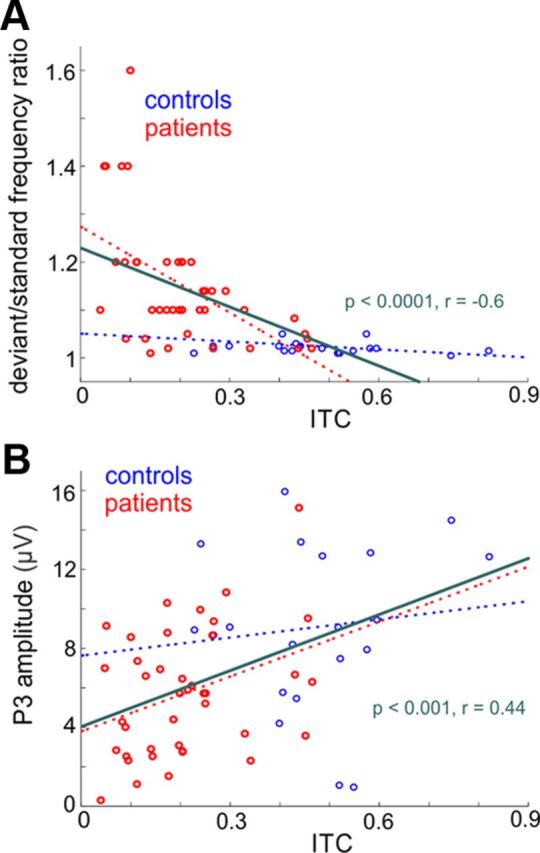
Recent neuroscience advances suggest that when interacting with our environment, along with previous experience, we use contextual cues and regularities to form predictions that guide our perceptions and actions. The goal of such active "predictive sensing" is to selectively enhance the processing and representation of behaviorally relevant information in an efficient manner. Since a hallmark of schizophrenia is impaired information selection, we tested whether this deficiency stems from dysfunctional predictive sensing by measuring the degree to which neuronal activity predicts relevant events. In healthy subjects, we established that these mechanisms are engaged in an effort-dependent manner and that, based on a correspondence between human scalp and intracranial nonhuman primate recordings, their main role is a predictive suppression of excitability in task-irrelevant regions. In contrast, schizophrenia patients displayed a reduced alignment of neuronal activity to attended stimuli, which correlated with their behavioral performance deficits and clinical symptoms. These results support the relevance of predictive sensing for normal and aberrant brain function, and highlight the importance of neuronal mechanisms that mold internal ongoing neuronal activity to model key features of the external environment.
Figures
Similar articles
-
Visuo-auditory interactions in the primary visual cortex of the behaving monkey: electrophysiological evidence.BMC Neurosci. 2008 Aug 12;9:79. doi: 10.1186/1471-2202-9-79. BMC Neurosci. 2008. PMID: 18699988 Free PMC article.
-
A predictive coding account of MMN reduction in schizophrenia.Biol Psychol. 2016 Apr;116:68-74. doi: 10.1016/j.biopsycho.2015.10.011. Epub 2015 Nov 12. Biol Psychol. 2016. PMID: 26582536
-
Occipital γ response to auditory stimulation in patients with schizophrenia.Int J Psychophysiol. 2011 Jan;79(1):3-8. doi: 10.1016/j.ijpsycho.2010.10.011. Epub 2010 Nov 5. Int J Psychophysiol. 2011. PMID: 21056599
-
The mismatch-negativity (MMN) component of the auditory event-related potential to violations of abstract regularities: a review.Int J Psychophysiol. 2013 May;88(2):109-23. doi: 10.1016/j.ijpsycho.2013.03.015. Epub 2013 Mar 28. Int J Psychophysiol. 2013. PMID: 23542165 Review.
-
Cortical Microcircuit Mechanisms of Mismatch Negativity and Its Underlying Subcomponents.Front Neural Circuits. 2020 Mar 31;14:13. doi: 10.3389/fncir.2020.00013. eCollection 2020. Front Neural Circuits. 2020. PMID: 32296311 Free PMC article. Review.
Cited by
-
Predictive Modulation of Corticospinal Excitability and Implicit Encoding of Movement Probability in Schizophrenia.Schizophr Bull. 2019 Oct 24;45(6):1358-1366. doi: 10.1093/schbul/sby186. Schizophr Bull. 2019. PMID: 30561714 Free PMC article.
-
Sensory and cross-network contributions to response inhibition in patients with schizophrenia.Neuroimage Clin. 2018 Jan 6;18:31-39. doi: 10.1016/j.nicl.2018.01.001. eCollection 2018. Neuroimage Clin. 2018. PMID: 29868440 Free PMC article.
-
Psychiatric Disorders and lncRNAs: A Synaptic Match.Int J Mol Sci. 2020 Apr 25;21(9):3030. doi: 10.3390/ijms21093030. Int J Mol Sci. 2020. PMID: 32344798 Free PMC article. Review.
-
Entrainment of neural oscillations as a modifiable substrate of attention.Trends Cogn Sci. 2014 Jun;18(6):300-9. doi: 10.1016/j.tics.2014.02.005. Epub 2014 Mar 12. Trends Cogn Sci. 2014. PMID: 24630166 Free PMC article. Review.
-
As without, so within: how the brain's temporo-spatial alignment to the environment shapes consciousness.Interface Focus. 2023 Apr 14;13(3):20220076. doi: 10.1098/rsfs.2022.0076. eCollection 2023 Jun 6. Interface Focus. 2023. PMID: 37065263 Free PMC article.
References
Publication types
MeSH terms
Grants and funding
- DC010415/DC/NIDCD NIH HHS/United States
- R01 MH049334/MH/NIMH NIH HHS/United States
- MH061989/MH/NIMH NIH HHS/United States
- MH082790/MH/NIMH NIH HHS/United States
- R01 DC011490/DC/NIDCD NIH HHS/United States
- MH086385/MH/NIMH NIH HHS/United States
- R37 MH049334/MH/NIMH NIH HHS/United States
- MH49334/MH/NIMH NIH HHS/United States
- R01 MH061989/MH/NIMH NIH HHS/United States
- P50 MH086385/MH/NIMH NIH HHS/United States
- DC012947/DC/NIDCD NIH HHS/United States
- R24 MH082790/MH/NIMH NIH HHS/United States
- R21 DC010415/DC/NIDCD NIH HHS/United States
- R01 DC012947/DC/NIDCD NIH HHS/United States
- R01 MH060358/MH/NIMH NIH HHS/United States
- MH060358/MH/NIMH NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical