

The endocytosis of cellulose synthase in Arabidopsis is dependent on μ2, a clathrin-mediated endocytosis adaptin
- PMID: 23843604
- PMCID: PMC3762637
- DOI: 10.1104/pp.113.221234
The endocytosis of cellulose synthase in Arabidopsis is dependent on μ2, a clathrin-mediated endocytosis adaptin
Abstract
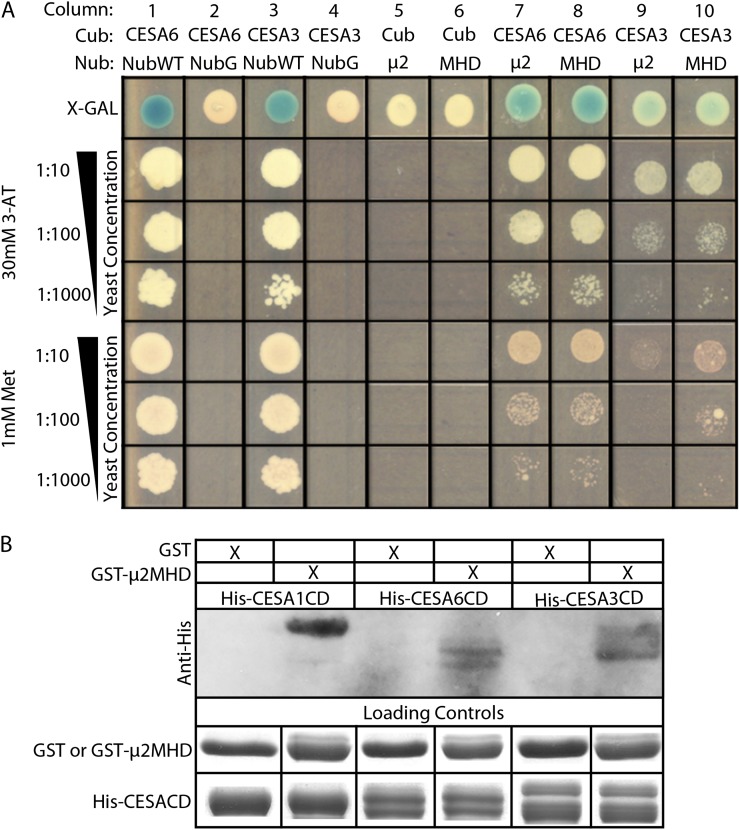
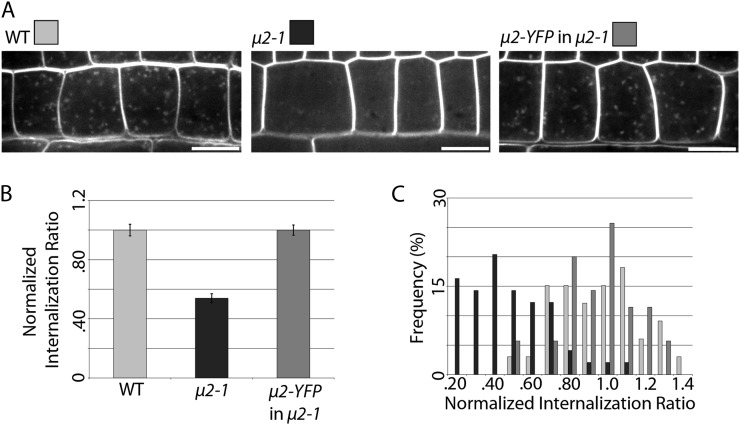
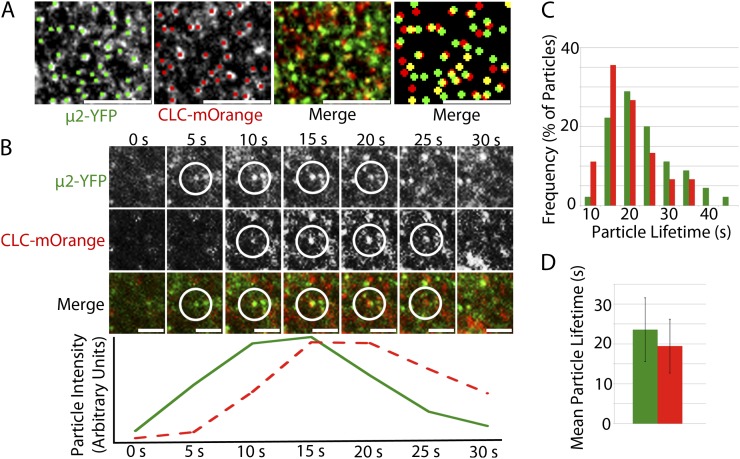
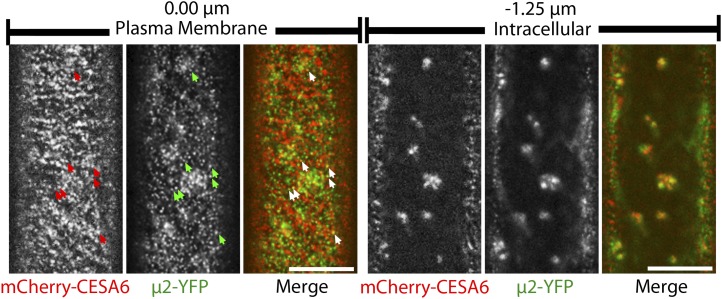
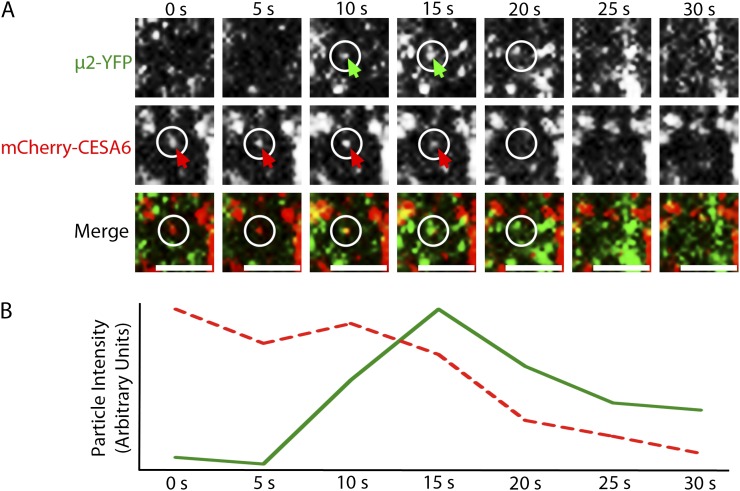
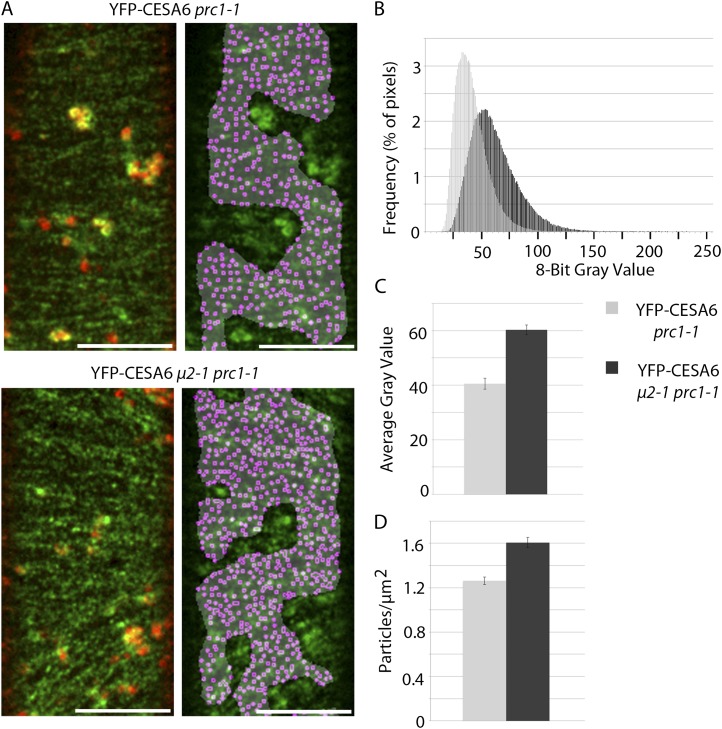
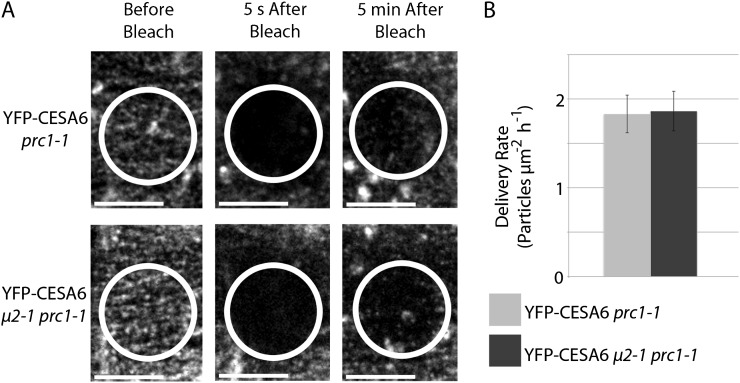
Clathrin-mediated endocytosis (CME) is the best-characterized type of endocytosis in eukaryotic cells. Plants appear to possess all of the molecular components necessary to carry out CME; however, functional characterization of the components is still in its infancy. A yeast two-hybrid screen identified μ2 as a putative interaction partner of CELLULOSE SYNTHASE6 (CESA6). Arabidopsis (Arabidopsis thaliana) μ2 is homologous to the medium subunit 2 of the mammalian ADAPTOR PROTEIN COMPLEX2 (AP2). In mammals, the AP2 complex acts as the central hub of CME by docking to the plasma membrane while concomitantly recruiting cargo proteins, clathrin triskelia, and accessory proteins to the sites of endocytosis. We confirmed that μ2 interacts with multiple CESA proteins through the μ-homology domain of μ2, which is involved in specific interactions with endocytic cargo proteins in mammals. Consistent with its role in mediating the endocytosis of cargos at the plasma membrane, μ2-YELLOW FLUORESCENT PROTEIN localized to transient foci at the plasma membrane, and loss of μ2 resulted in defects in bulk endocytosis. Furthermore, loss of μ2 led to increased accumulation of YELLOW FLUORESCENT PROTEIN-CESA6 particles at the plasma membrane. Our results suggest that CESA represents a new class of CME cargo proteins and that plant cells might regulate cellulose synthesis by controlling the abundance of active CESA complexes at the plasma membrane through CME.
Figures
References
-
- Alonso JM, Stepanova AN, Leisse TJ, Kim CJ, Chen HM, Shinn P, Stevenson DK, Zimmerman J, Barajas P, Cheuk R, et al. (2003) Genome-wide insertional mutagenesis of Arabidopsis thaliana. Science 301: 653–657 - PubMed
-
- Baluska F, Liners F, Hlavacka A, Schlicht M, Van Cutsem P, McCurdy DW, Menzel D. (2005) Cell wall pectins and xyloglucans are internalized into dividing root cells and accumulate within cell plates during cytokinesis. Protoplasma 225: 141–155 - PubMed
-
- Bandmann V, Homann U. (2012) Clathrin-independent endocytosis contributes to uptake of glucose into BY-2 protoplasts. Plant J 70: 578–584 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
