

Signaling inputs to invadopodia and podosomes
- PMID: 23843616
- PMCID: PMC3711196
- DOI: 10.1242/jcs.079475
Signaling inputs to invadopodia and podosomes
Abstract
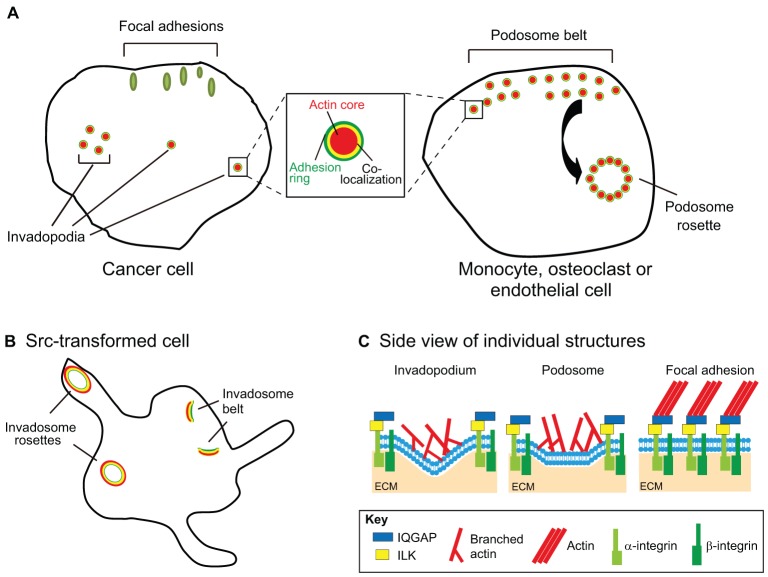
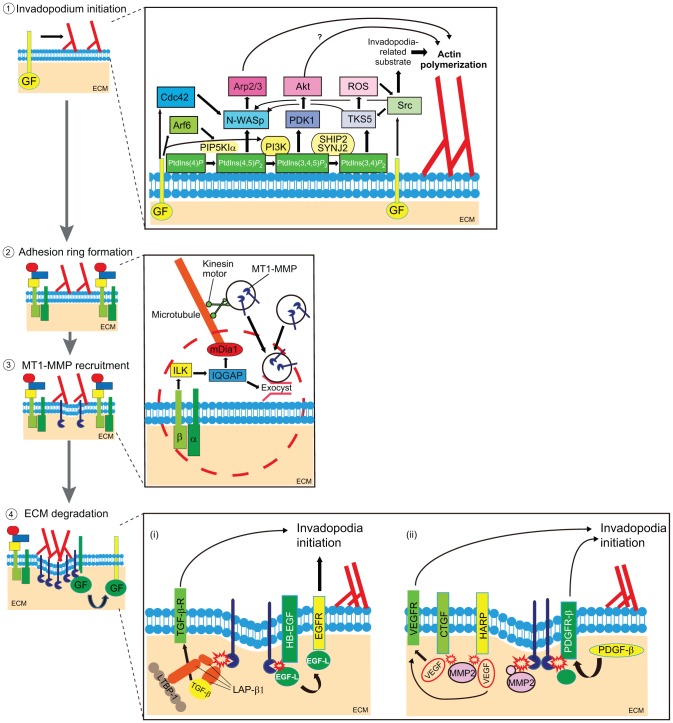
Remodeling of extracellular matrix (ECM) is a fundamental cell property that allows cells to alter their microenvironment and move through tissues. Invadopodia and podosomes are subcellular actin-rich structures that are specialized for matrix degradation and are formed by cancer and normal cells, respectively. Although initial studies focused on defining the core machinery of these two structures, recent studies have identified inputs from both growth factor and adhesion signaling as crucial for invasive activity. This Commentary will outline the current knowledge on the upstream signaling inputs to invadopodia and podosomes and their role in governing distinct stages of these invasive structures. We discuss invadopodia and podosomes as adhesion structures and highlight new data showing that invadopodia-associated adhesion rings promote the maturation of already-formed invadopodia. We present a model in which growth factor stimulation leads to phosphoinositide 3-kinase (PI3K) activity and formation of invadopodia, whereas adhesion signaling promotes exocytosis of proteinases at invadopodia.
Keywords: Actin cytoskeleton; Adhesion; Growth factor; Invadopodia; PI3K; Podosome; phosphoinositide 3-kinase.
Figures
References
-
- Artym V. V., Zhang Y., Seillier-Moiseiwitsch F., Yamada K. M., Mueller S. C. (2006). Dynamic interactions of cortactin and membrane type 1 matrix metalloproteinase at invadopodia: defining the stages of invadopodia formation and function. Cancer Res. 66, 3034–3043 10.1158/0008-5472.CAN-05-2177 - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
