

Phosphorylation of serine 4,642 in the C-terminus of plectin by MNK2 and PKA modulates its interaction with intermediate filaments
- PMID: 23843618
- PMCID: PMC3772390
- DOI: 10.1242/jcs.127779
Phosphorylation of serine 4,642 in the C-terminus of plectin by MNK2 and PKA modulates its interaction with intermediate filaments
Abstract
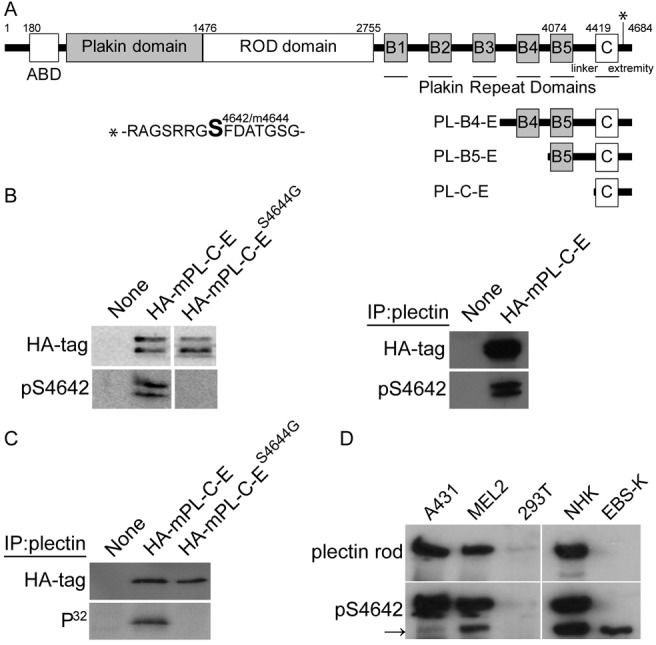
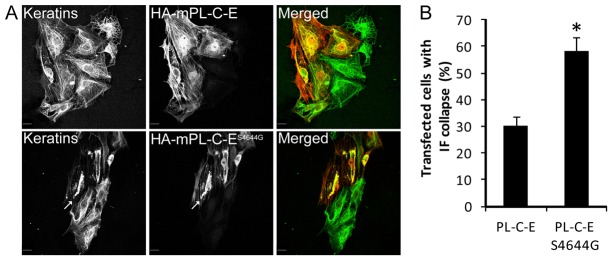
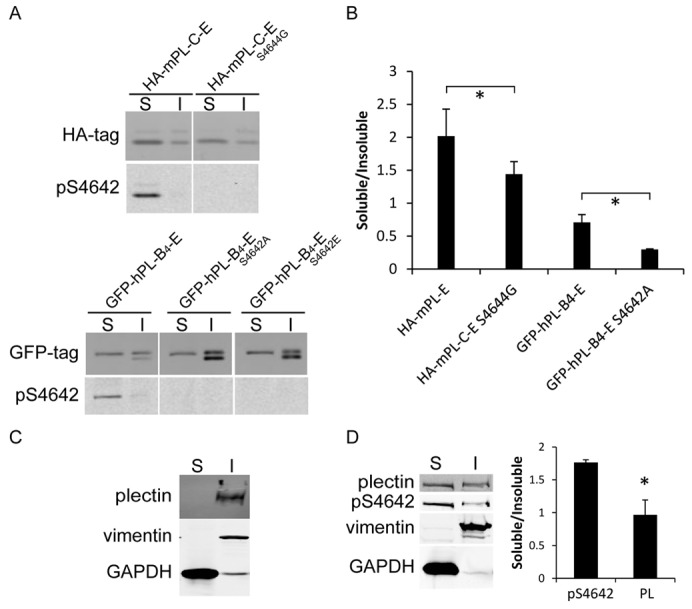
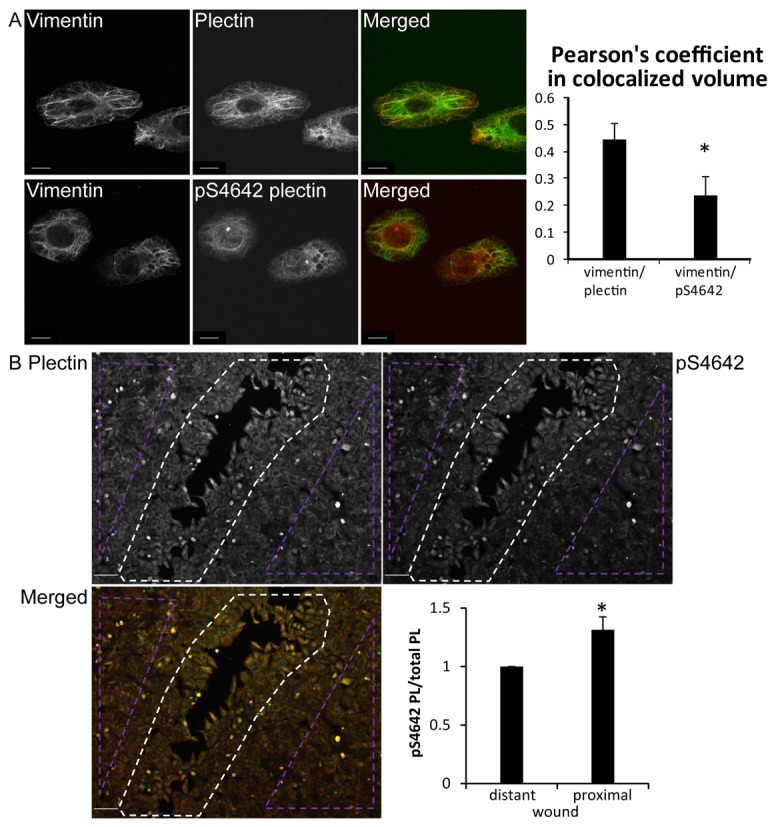
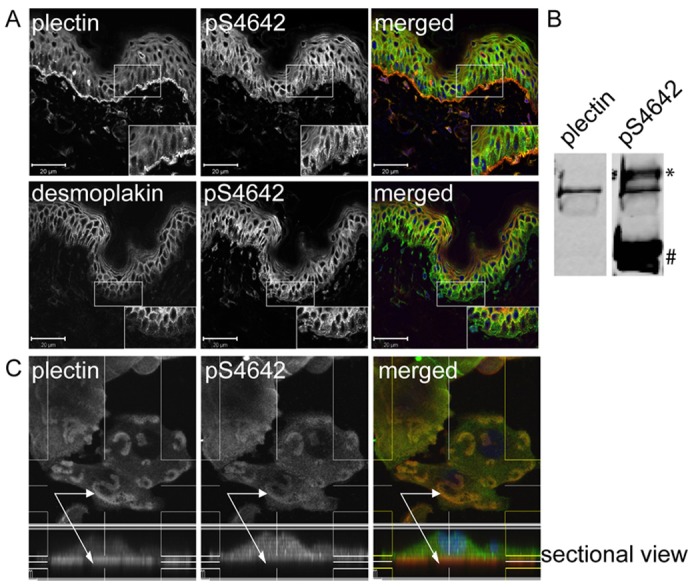
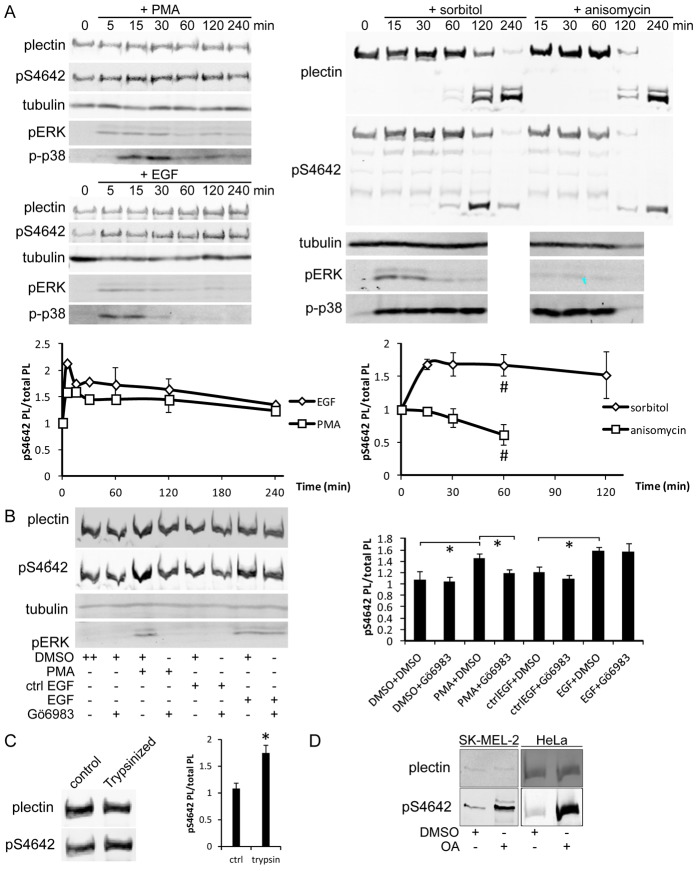
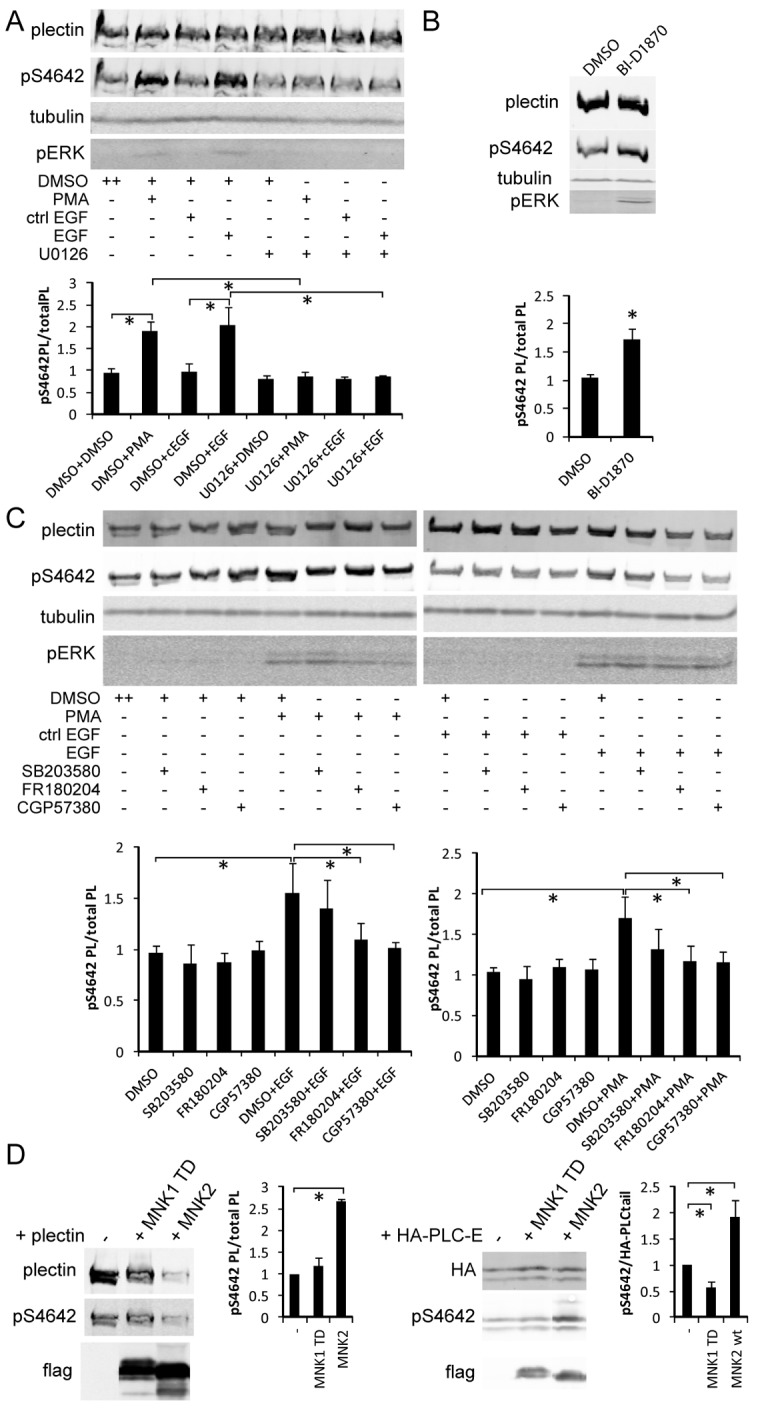
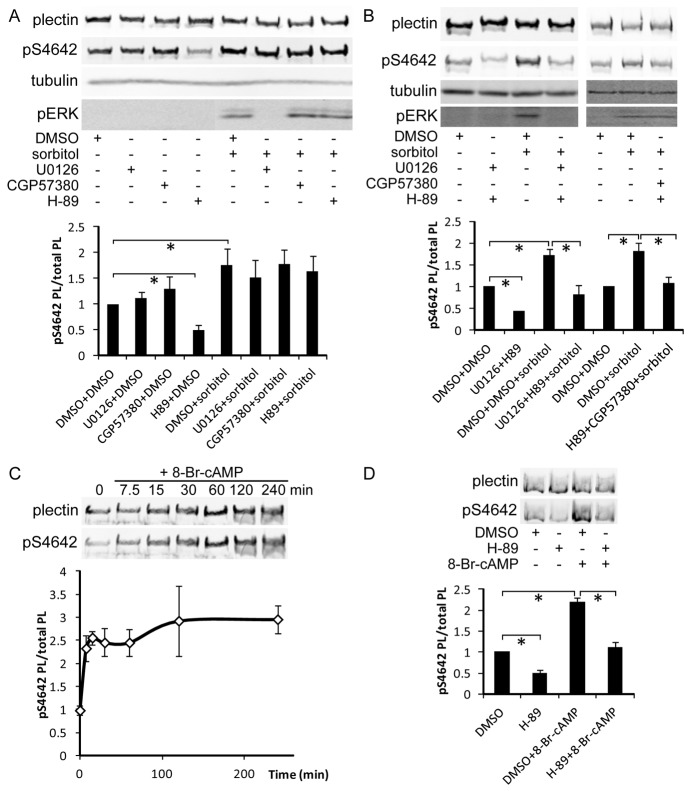
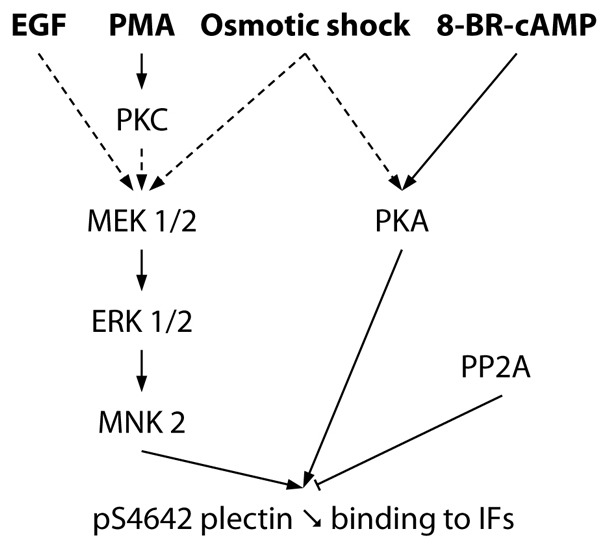
Plectin is a versatile cytolinker of the plakin family conferring cell resilience to mechanical stress in stratified epithelia and muscles. It acts as a critical organizer of the cytoskeletal system by tethering various intermediate filament (IF) networks through its C-terminal IF-binding domain (IFBD). Mutations affecting the IFBD cause devastating human diseases. Here, we show that serine 4642, which is located in the extreme C-terminus of plectin, is phosphorylated in different cell lines. Phosphorylation of S4642 decreased the ability of plectin IFBD to associate with various IFs, as assessed by immunofluorescence microscopy and cell fractionation studies, as well as in yeast two-hybrid assays. Plectin phosphorylated at S4642 was reduced at sites of IF network anchorage along cell-substrate contacts in both skin and cultured keratinocytes. Treatment of SK-MEL-2 and HeLa cells with okadaic acid increased plectin S4642 phosphorylation, suggesting that protein phosphatase 2A dephosphorylates this residue. Moreover, plectin S4642 phosphorylation was enhanced after cell treatment with EGF, phorbol ester, sorbitol and 8-bromo-cyclic AMP, as well as during wound healing and protease-mediated cell detachment. Using selective protein kinase inhibitors, we identified two different kinases that modulate the phosphorylation of plectin S4642 in HeLa cells: MNK2, which is downstream of the ERK1/2-dependent MAPK cascade, and PKA. Our study indicates that phosphorylation of S4642 has an important regulatory role in the interaction of plectin with IFs and identifies a novel link between MNK2 and the cytoskeleton.
Keywords: Cytoskeleton; Intermediate filaments; Plakin; Plectin; Protein phosphorylation.
Figures
Similar articles
-
Interaction of plectin and intermediate filaments.J Dermatol Sci. 2012 Apr;66(1):44-50. doi: 10.1016/j.jdermsci.2012.01.008. Epub 2012 Jan 24. J Dermatol Sci. 2012. PMID: 22333906
-
Plectin-intermediate filament partnership in skin, skeletal muscle, and peripheral nerve.Histochem Cell Biol. 2013 Jul;140(1):33-53. doi: 10.1007/s00418-013-1102-0. Epub 2013 Jun 9. Histochem Cell Biol. 2013. PMID: 23748243 Free PMC article. Review.
-
Plectin-controlled keratin cytoarchitecture affects MAP kinases involved in cellular stress response and migration.J Cell Biol. 2006 Aug 14;174(4):557-68. doi: 10.1083/jcb.200605172. J Cell Biol. 2006. PMID: 16908671 Free PMC article.
-
Dose-dependent linkage, assembly inhibition and disassembly of vimentin and cytokeratin 5/14 filaments through plectin's intermediate filament-binding domain.J Cell Sci. 2000 Feb;113 ( Pt 3):483-91. doi: 10.1242/jcs.113.3.483. J Cell Sci. 2000. PMID: 10639335
-
Networking and anchoring through plectin: a key to IF functionality and mechanotransduction.Curr Opin Cell Biol. 2015 Feb;32:21-9. doi: 10.1016/j.ceb.2014.10.002. Epub 2014 Oct 29. Curr Opin Cell Biol. 2015. PMID: 25460778 Review.
Cited by
-
GSK3- and PRMT-1-dependent modifications of desmoplakin control desmoplakin-cytoskeleton dynamics.J Cell Biol. 2015 Mar 2;208(5):597-612. doi: 10.1083/jcb.201406020. J Cell Biol. 2015. PMID: 25733715 Free PMC article.
-
Mnk kinase pathway: Cellular functions and biological outcomes.World J Biol Chem. 2014 Aug 26;5(3):321-33. doi: 10.4331/wjbc.v5.i3.321. World J Biol Chem. 2014. PMID: 25225600 Free PMC article. Review.
-
Plakins, a versatile family of cytolinkers: roles in skin integrity and in human diseases.J Invest Dermatol. 2014 Apr;134(4):885-894. doi: 10.1038/jid.2013.498. Epub 2013 Dec 19. J Invest Dermatol. 2014. PMID: 24352042 Review.
-
Regulation of intestinal epithelial intercellular adhesion and barrier function by desmosomal cadherin desmocollin-2.Mol Biol Cell. 2021 Apr 15;32(8):753-768. doi: 10.1091/mbc.E20-12-0775. Epub 2021 Feb 17. Mol Biol Cell. 2021. PMID: 33596089 Free PMC article.
-
Proteomics of Plasma and Plasma-Treated Podocytes: Application to Focal and Segmental Glomerulosclerosis.Int J Mol Sci. 2023 Jul 28;24(15):12124. doi: 10.3390/ijms241512124. Int J Mol Sci. 2023. PMID: 37569500 Free PMC article.
References
-
- Bagowski C. P., Besser J., Frey C. R., Ferrell J. E., Jr (2003). The JNK cascade as a biochemical switch in mammalian cells: ultrasensitive and all-or-none responses. Curr. Biol. 13, 315–320 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
