

Iduronic acid in chondroitin/dermatan sulfate affects directional migration of aortic smooth muscle cells
- PMID: 23843960
- PMCID: PMC3699603
- DOI: 10.1371/journal.pone.0066704
Iduronic acid in chondroitin/dermatan sulfate affects directional migration of aortic smooth muscle cells
Abstract
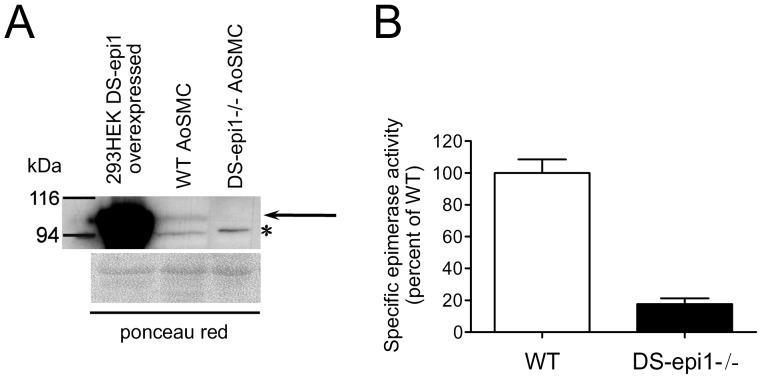
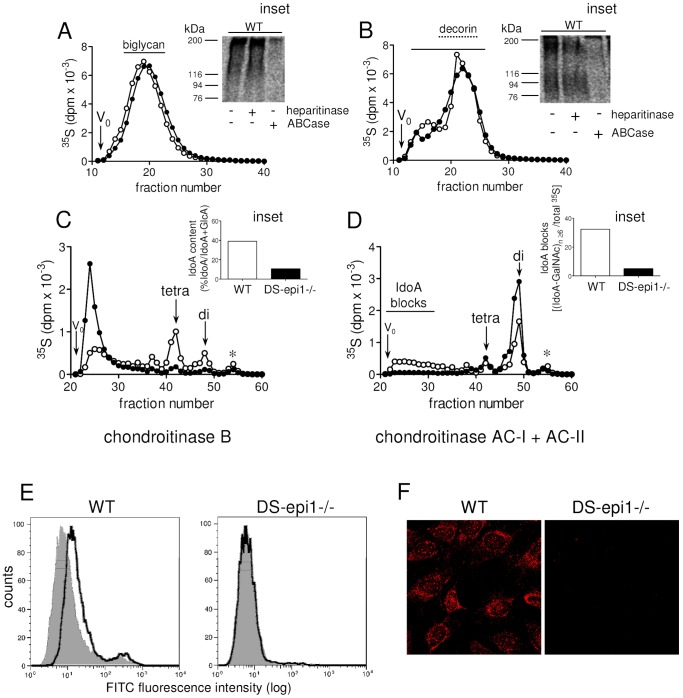
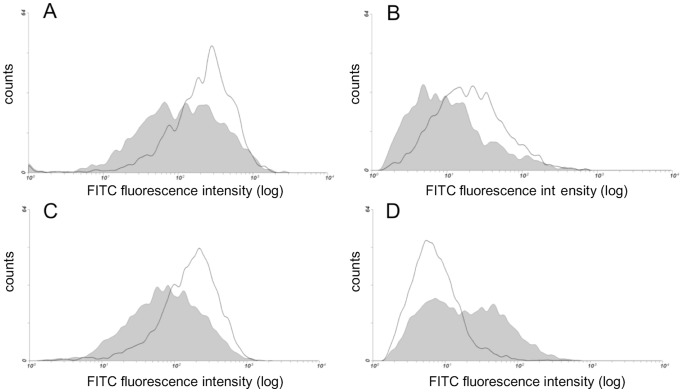
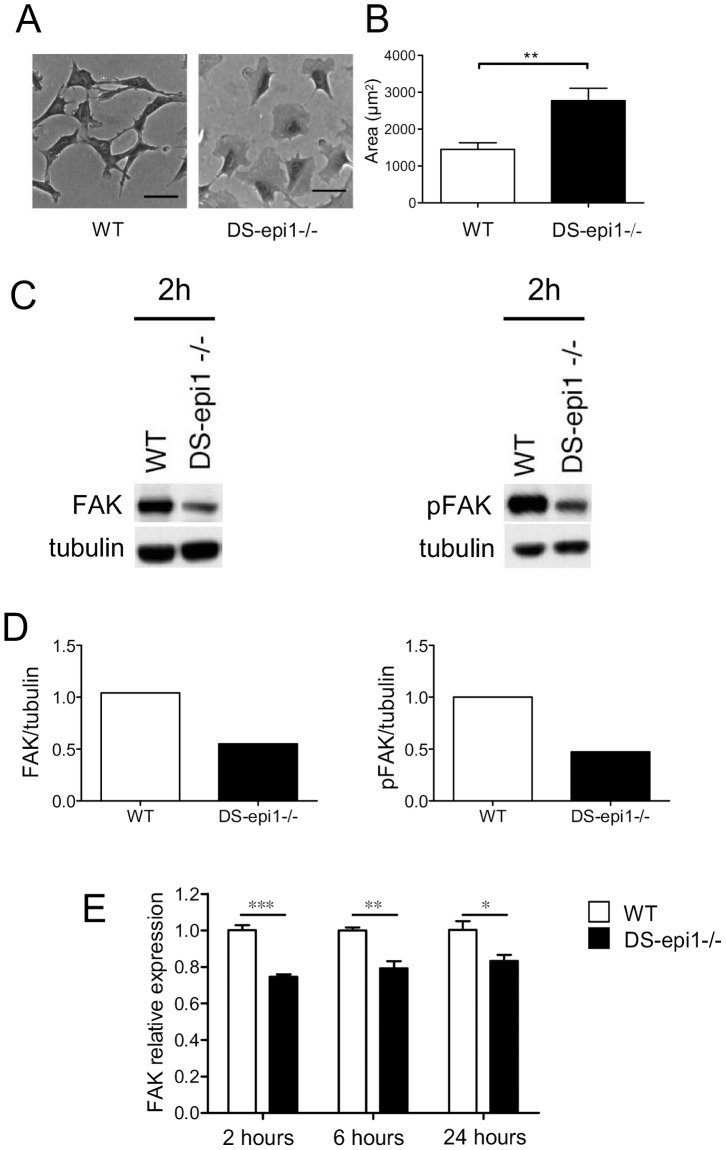
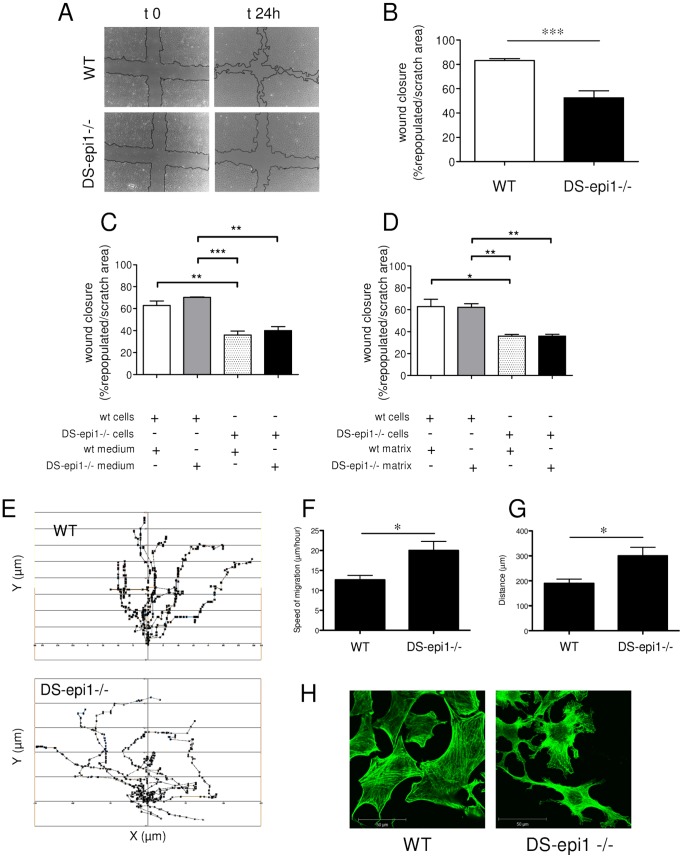
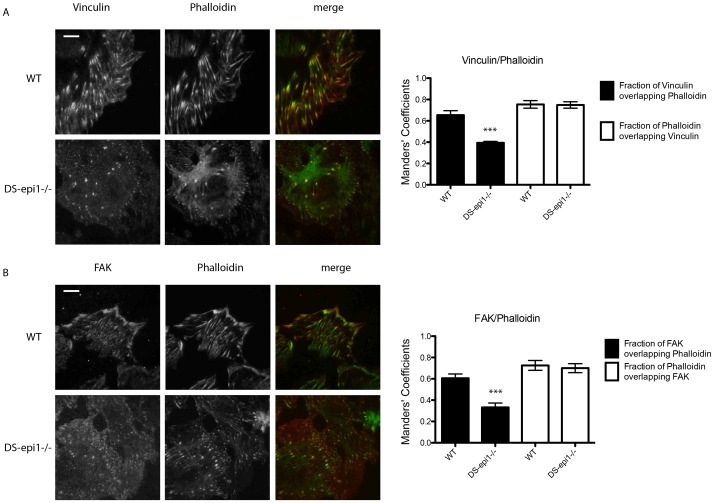
Aortic smooth muscle cells produce chondroitin/dermatan sulfate (CS/DS) proteoglycans that regulate extracellular matrix organization and cell behavior in normal and pathological conditions. A unique feature of CS/DS proteoglycans is the presence of iduronic acid (IdoA), catalyzed by two DS epimerases. Functional ablation of DS-epi1, the main epimerase in these cells, resulted in a major reduction of IdoA both on cell surface and in secreted CS/DS proteoglycans. Downregulation of IdoA led to delayed ability to re-populate wounded areas due to loss of directional persistence of migration. DS-epi1-/- aortic smooth muscle cells, however, had not lost the general property of migration showing even increased speed of movement compared to wild type cells. Where the cell membrane adheres to the substratum, stress fibers were denser whereas focal adhesion sites were fewer. Total cellular expression of focal adhesion kinase (FAK) and phospho-FAK (pFAK) was decreased in mutant cells compared to control cells. As many pathological conditions are dependent on migration, modulation of IdoA content may point to therapeutic strategies for diseases such as cancer and atherosclerosis.
Conflict of interest statement
Figures
Similar articles
-
Dermatan sulfate is involved in the tumorigenic properties of esophagus squamous cell carcinoma.Cancer Res. 2012 Apr 15;72(8):1943-52. doi: 10.1158/0008-5472.CAN-11-1351. Epub 2012 Feb 20. Cancer Res. 2012. PMID: 22350411 Free PMC article.
-
Dermatan sulfate epimerase 1-deficient mice have reduced content and changed distribution of iduronic acids in dermatan sulfate and an altered collagen structure in skin.Mol Cell Biol. 2009 Oct;29(20):5517-28. doi: 10.1128/MCB.00430-09. Epub 2009 Aug 17. Mol Cell Biol. 2009. PMID: 19687302 Free PMC article.
-
Dermatan sulfate epimerase 2 is the predominant isozyme in the formation of the chondroitin sulfate/dermatan sulfate hybrid structure in postnatal developing mouse brain.Glycobiology. 2011 May;21(5):565-74. doi: 10.1093/glycob/cwq208. Epub 2010 Dec 21. Glycobiology. 2011. PMID: 21177331
-
Iduronic acid in chondroitin/dermatan sulfate: biosynthesis and biological function.J Histochem Cytochem. 2012 Dec;60(12):916-25. doi: 10.1369/0022155412459857. Epub 2012 Aug 16. J Histochem Cytochem. 2012. PMID: 22899863 Free PMC article. Review.
-
Biological functions of iduronic acid in chondroitin/dermatan sulfate.FEBS J. 2013 May;280(10):2431-46. doi: 10.1111/febs.12214. Epub 2013 Mar 28. FEBS J. 2013. PMID: 23441919 Free PMC article. Review.
Cited by
-
Melanoma Cell Adhesion and Migration Is Modulated by the Uronyl 2-O Sulfotransferase.PLoS One. 2017 Jan 20;12(1):e0170054. doi: 10.1371/journal.pone.0170054. eCollection 2017. PLoS One. 2017. PMID: 28107390 Free PMC article.
-
Iduronate-2-Sulfatase-Regulated Dermatan Sulfate Levels Potentiate the Invasion of Breast Cancer Epithelia through Collagen Matrix.J Clin Med. 2019 Sep 30;8(10):1562. doi: 10.3390/jcm8101562. J Clin Med. 2019. PMID: 31574977 Free PMC article.
-
An Overview of in vivo Functions of Chondroitin Sulfate and Dermatan Sulfate Revealed by Their Deficient Mice.Front Cell Dev Biol. 2021 Nov 24;9:764781. doi: 10.3389/fcell.2021.764781. eCollection 2021. Front Cell Dev Biol. 2021. PMID: 34901009 Free PMC article. Review.
-
Human genetic disorders and knockout mice deficient in glycosaminoglycan.Biomed Res Int. 2014;2014:495764. doi: 10.1155/2014/495764. Epub 2014 Jul 13. Biomed Res Int. 2014. PMID: 25126564 Free PMC article. Review.
-
The structure of human dermatan sulfate epimerase 1 emphasizes the importance of C5-epimerization of glucuronic acid in higher organisms.Chem Sci. 2020 Dec 8;12(5):1869-1885. doi: 10.1039/d0sc05971d. eCollection 2021 Feb 7. Chem Sci. 2020. PMID: 33815739 Free PMC article.
References
-
- Couchman JR (2010) Transmembrane signaling proteoglycans. Annu Rev Cell Dev Biol 26: 89–114. - PubMed
-
- Woods A, Couchman JR (2000) Integrin modulation by lateral association. J Biol Chem 275: 24233–24236. - PubMed
-
- Maccarana M, Olander B, Malmstrom J, Tiedemann K, Aebersold R, et al. (2006) Biosynthesis of dermatan sulfate: chondroitin-glucuronate C5-epimerase is identical to SART2. J Biol Chem 281: 11560–11568. - PubMed
-
- Ilarraza Lomeli H, Herrera Franco R, Lomeli Rivas A, Zavala Ramirez J, Martinez Ramirez L, et al. (2009) [National Registry of Cardiac Rehabilitation Programs in Mexico]. Arch Cardiol Mex 79: 63–72. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
