

Shear stress regulates late EPC differentiation via mechanosensitive molecule-mediated cytoskeletal rearrangement
- PMID: 23844056
- PMCID: PMC3699607
- DOI: 10.1371/journal.pone.0067675
Shear stress regulates late EPC differentiation via mechanosensitive molecule-mediated cytoskeletal rearrangement
Abstract
Background: Previous studies have demonstrated that endothelial progenitor cells (EPCs), in particular late EPCs, play important roles in endothelial maintenance and repair. Recent evidence has revealed shear stress as a key regulator for EPC differentiation. However, the underlying mechanisms regulating the shear stress-induced EPC differentiation have not been understood completely. The present study was undertaken to further investigate the effects of shear stress on the late EPC differentiation, and to elucidate the signal mechanism involved.
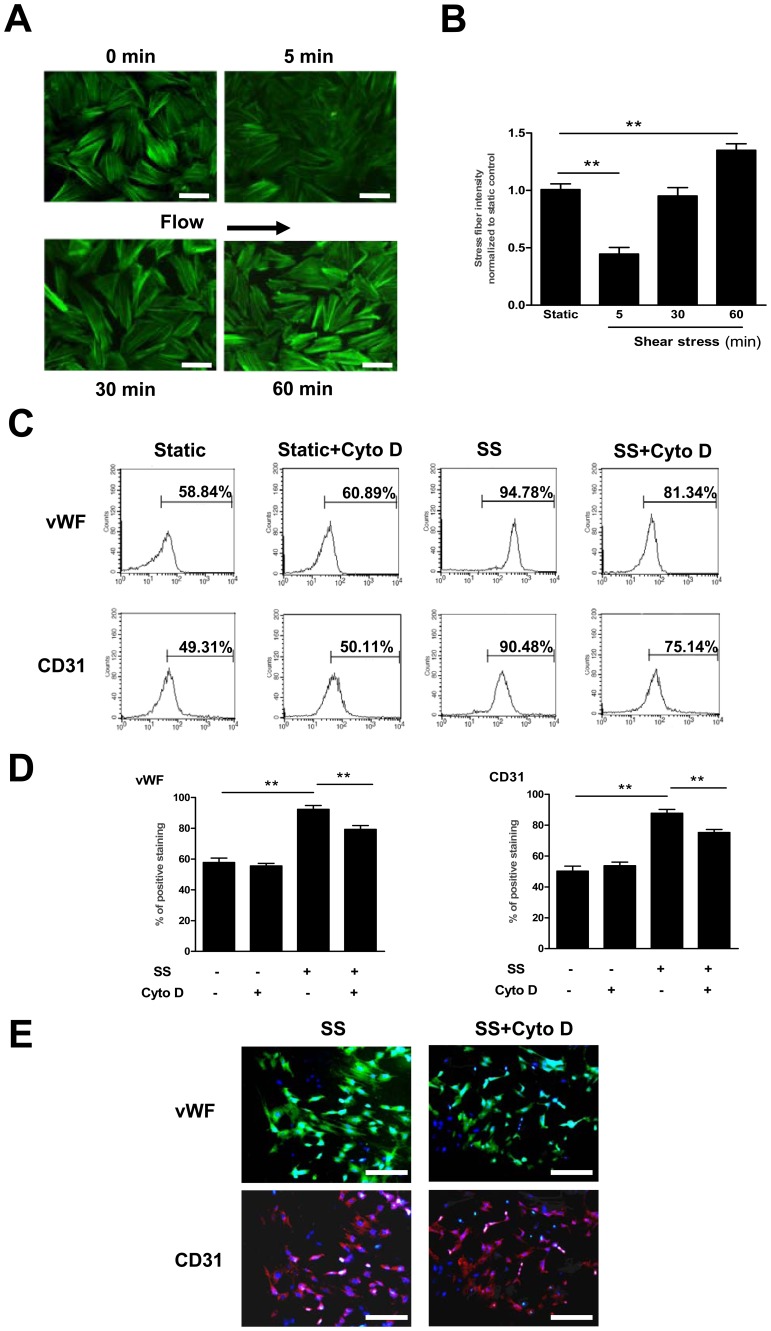
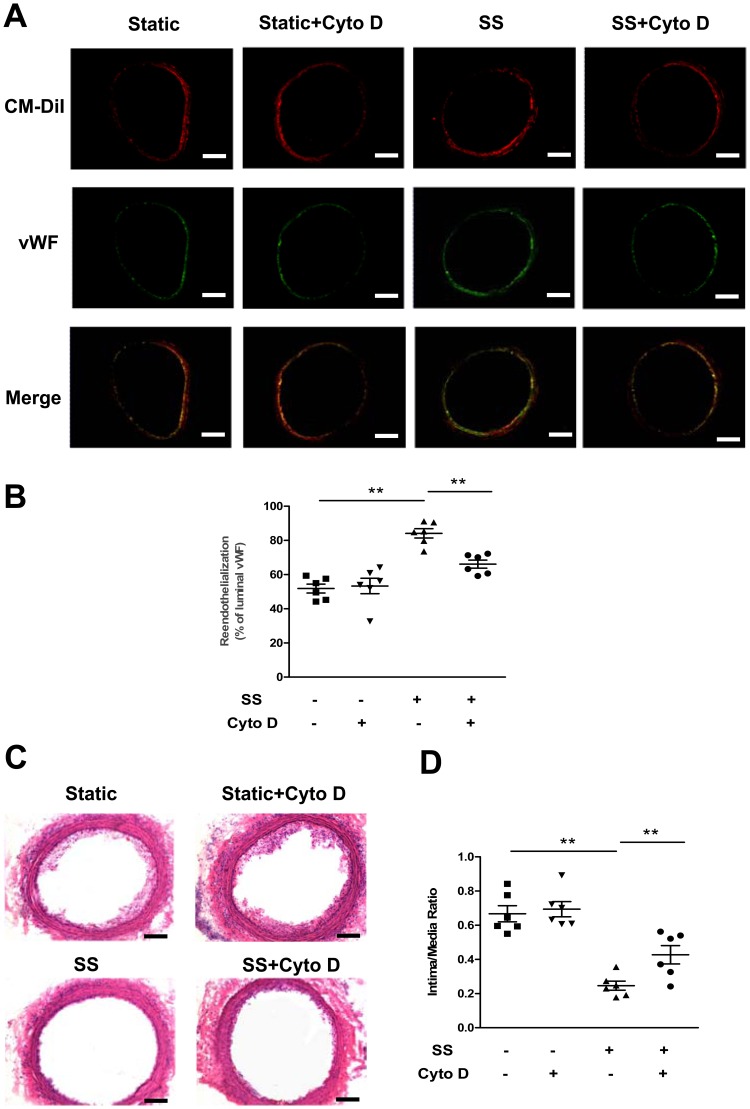
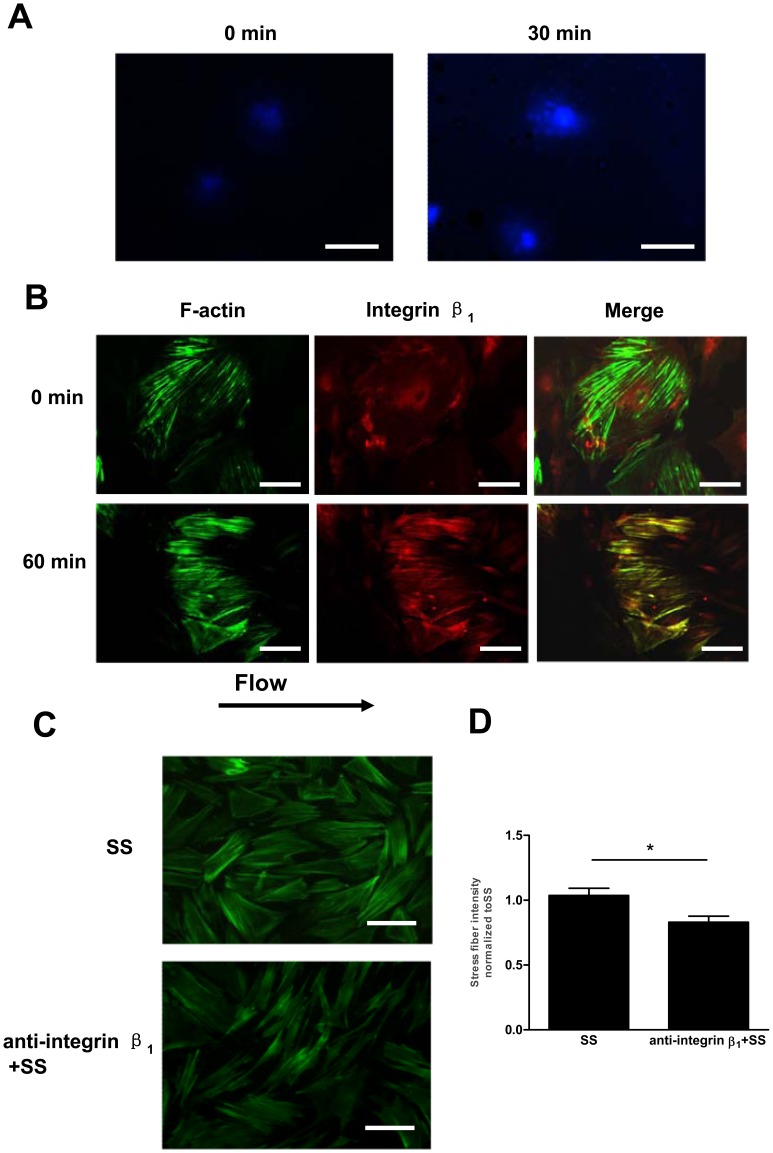
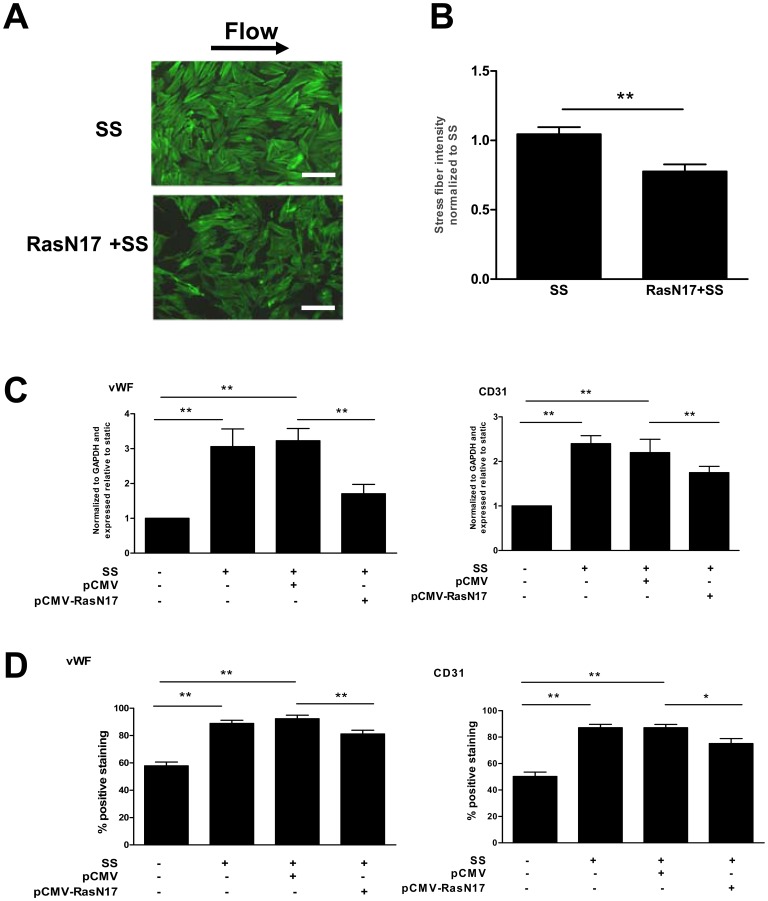
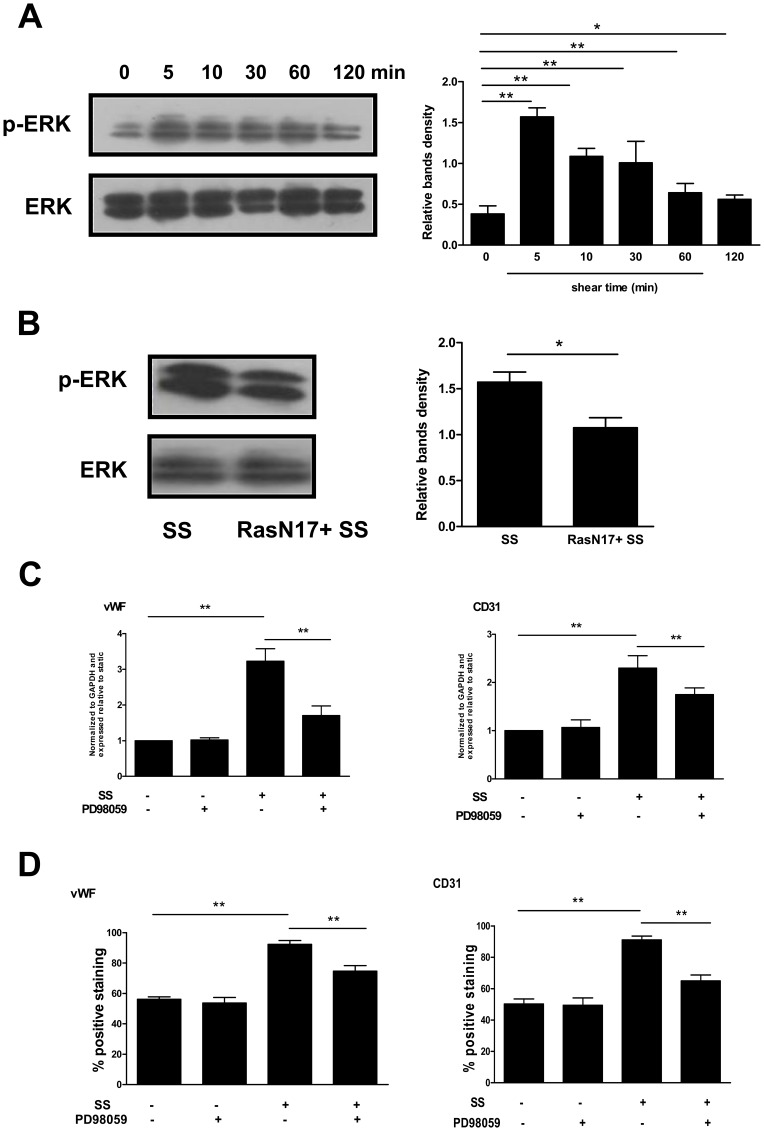
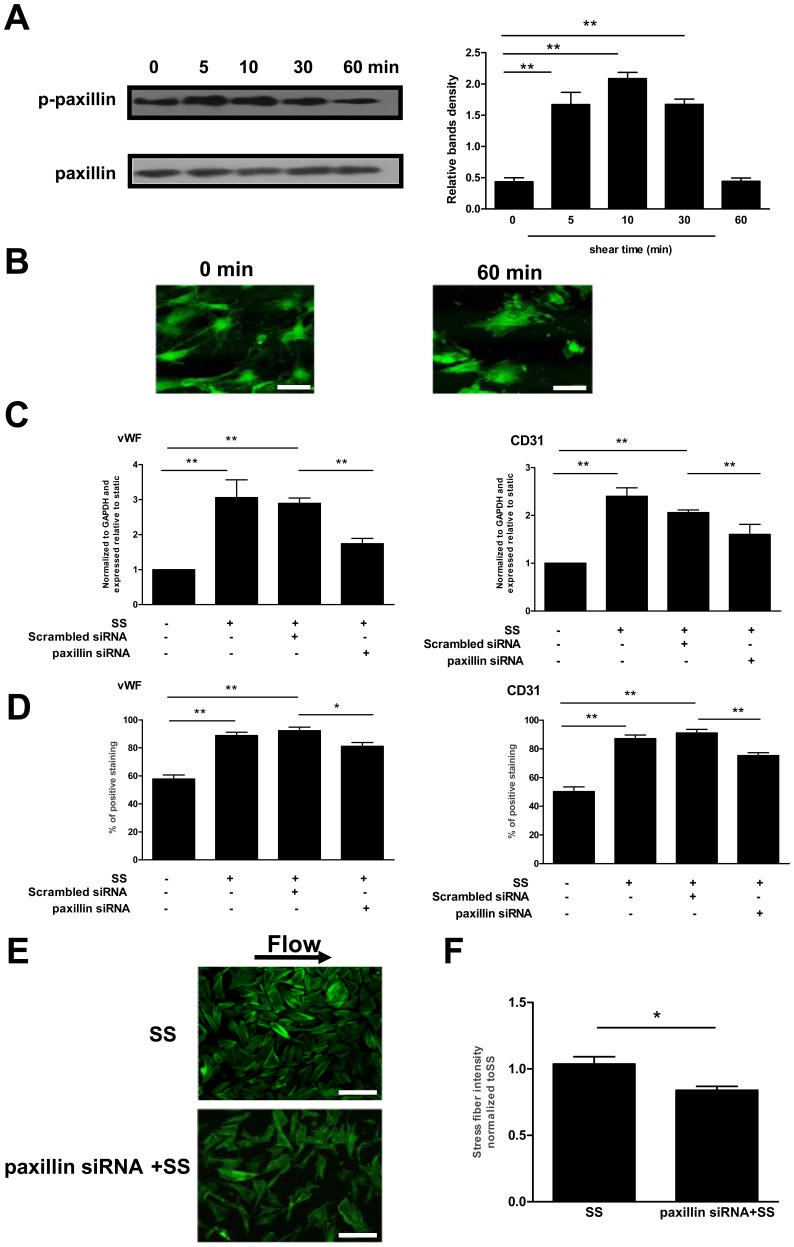
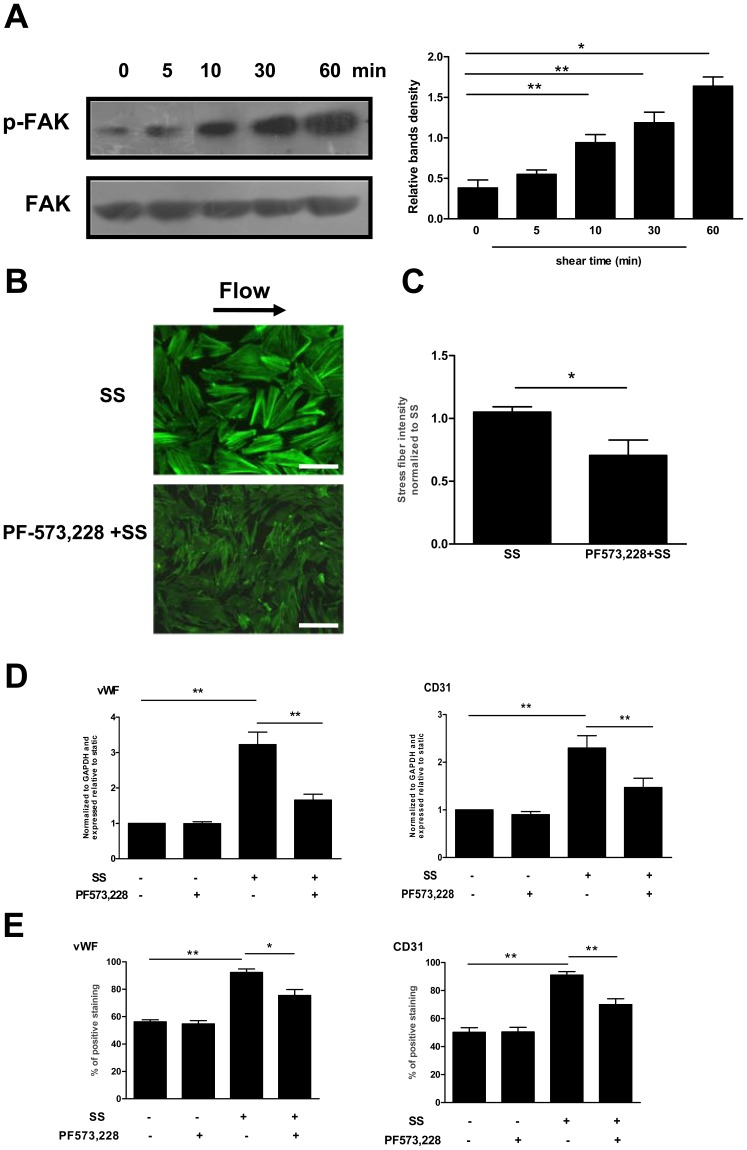
Methodology/principal finding: In vitro and in vivo assays revealed that cytoskeletal remodeling was involved in the shear stress-upregulated expression of endothelial markers vWF and CD31 in late EPCs, with subsequently increased in vivo reendothelialization after arterial injury. Moreover, shear stress activated several mechanosensitive molecules including integrin β1, Ras, ERK1/2, paxillin and FAK, which were all involved in both cytoskeletal rearrangement and cell differentiation in response to shear stress in late EPCs.
Conclusions/significance: Shear stress is a key regulator for late EPC differentiation into endothelial cells, which is important for vascular repair, and the cytoskeletal rearrangement mediated by the activation of the cascade of integrin β1, Ras, ERK1/2, paxillin and FAK is crucial in this process.
Conflict of interest statement
Figures
Similar articles
-
Function of Krüppel‑like factor 2 in the shear stress‑induced cell differentiation of endothelial progenitor cells to endothelial cells.Mol Med Rep. 2019 Mar;19(3):1739-1746. doi: 10.3892/mmr.2019.9819. Epub 2019 Jan 3. Mol Med Rep. 2019. PMID: 30628700
-
Shear stress augments the endothelial cell differentiation marker expression in late EPCs by upregulating integrins.Biochem Biophys Res Commun. 2012 Aug 24;425(2):419-25. doi: 10.1016/j.bbrc.2012.07.115. Epub 2012 Jul 27. Biochem Biophys Res Commun. 2012. PMID: 22846566
-
Shear stress and vascular smooth muscle cells promote endothelial differentiation of endothelial progenitor cells via activation of Akt.Clin Biomech (Bristol). 2008;23 Suppl 1:S118-24. doi: 10.1016/j.clinbiomech.2007.08.018. Epub 2007 Oct 24. Clin Biomech (Bristol). 2008. PMID: 17928113
-
Mechanosensitive microRNAs-role in endothelial responses to shear stress and redox state.Free Radic Biol Med. 2013 Sep;64:61-8. doi: 10.1016/j.freeradbiomed.2013.05.034. Epub 2013 May 30. Free Radic Biol Med. 2013. PMID: 23727269 Free PMC article. Review.
-
Mechanotransduction in endothelial cells: temporal signaling events in response to shear stress.J Vasc Res. 1997 May-Jun;34(3):212-9. doi: 10.1159/000159225. J Vasc Res. 1997. PMID: 9226303 Review.
Cited by
-
Influence of airway wall compliance on epithelial cell injury and adhesion during interfacial flows.J Appl Physiol (1985). 2014 Dec 1;117(11):1231-42. doi: 10.1152/japplphysiol.00752.2013. Epub 2014 Sep 11. J Appl Physiol (1985). 2014. PMID: 25213636 Free PMC article.
-
IFN-I Mediates Dysfunction of Endothelial Progenitor Cells in Atherosclerosis of Systemic Lupus Erythematosus.Front Immunol. 2020 Nov 11;11:581385. doi: 10.3389/fimmu.2020.581385. eCollection 2020. Front Immunol. 2020. PMID: 33262760 Free PMC article. Review.
-
Whole-Transcriptome Sequencing: a Powerful Tool for Vascular Tissue Engineering and Endothelial Mechanobiology.High Throughput. 2018 Feb 21;7(1):5. doi: 10.3390/ht7010005. High Throughput. 2018. PMID: 29485616 Free PMC article. Review.
-
Uremic Toxicity of Advanced Glycation End Products in CKD.J Am Soc Nephrol. 2016 Feb;27(2):354-70. doi: 10.1681/ASN.2014101047. Epub 2015 Aug 26. J Am Soc Nephrol. 2016. PMID: 26311460 Free PMC article. Review.
-
Effects of shear stress on differentiation of stem cells into endothelial cells.World J Stem Cells. 2021 Jul 26;13(7):894-913. doi: 10.4252/wjsc.v13.i7.894. World J Stem Cells. 2021. PMID: 34367483 Free PMC article. Review.
References
-
- Landmesser U, Hornig B, Drexler H (2004) Endothelial function: a critical determinant in atherosclerosis? Circulation 109: II27–33. - PubMed
-
- Deanfield JE, Halcox JP, Rabelink TJ (2007) Endothelial function and dysfunction: testing and clinical relevance. Circulation 115: 1285–1295. - PubMed
-
- Xu Q (2006) The impact of progenitor cells in atherosclerosis. Nat Clin Pract Cardiovasc Med 3: 94–101. - PubMed
-
- Werner N, Priller J, Laufs U, Endres M, Bohm M, et al. (2002) Bone marrow-derived progenitor cells modulate vascular reendothelialization and neointimal formation: effect of 3-hydroxy-3-methylglutaryl coenzyme a reductase inhibition. Arterioscler Thromb Vasc Biol 22: 1567–1572. - PubMed
-
- Tanaka K, Sata M, Natori T, Kim-Kaneyama JR, Nose K, et al. (2008) Circulating progenitor cells contribute to neointimal formation in nonirradiated chimeric mice. FASEB J 22: 428–436. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
