

Evolution and allometry of calcaneal elongation in living and extinct primates
- PMID: 23844094
- PMCID: PMC3701013
- DOI: 10.1371/journal.pone.0067792
Evolution and allometry of calcaneal elongation in living and extinct primates
Erratum in
- PLoS One. 2013;8(9). doi:10.1371/annotation/77784e61-3a43-4615-85bd-b6b4369568fd
Abstract
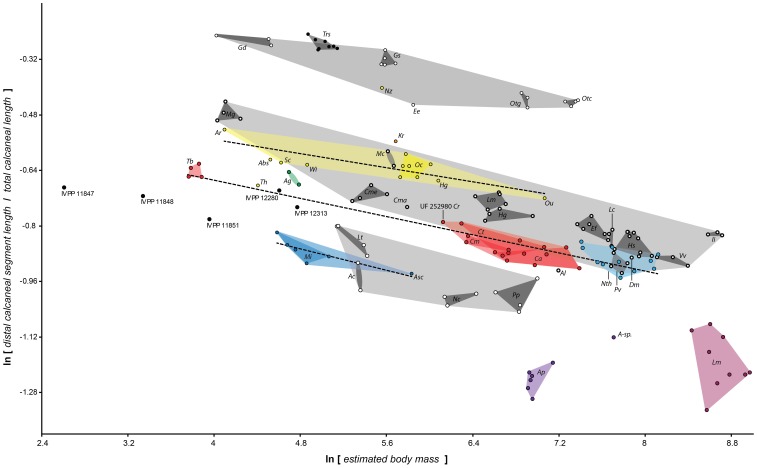
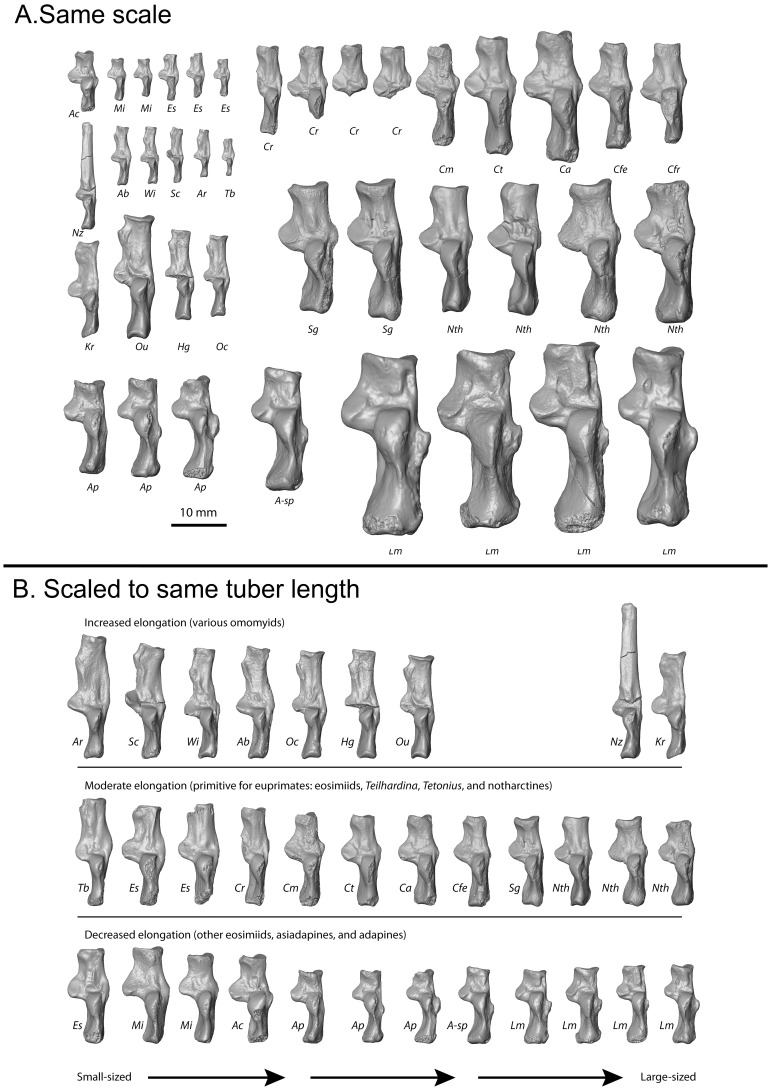
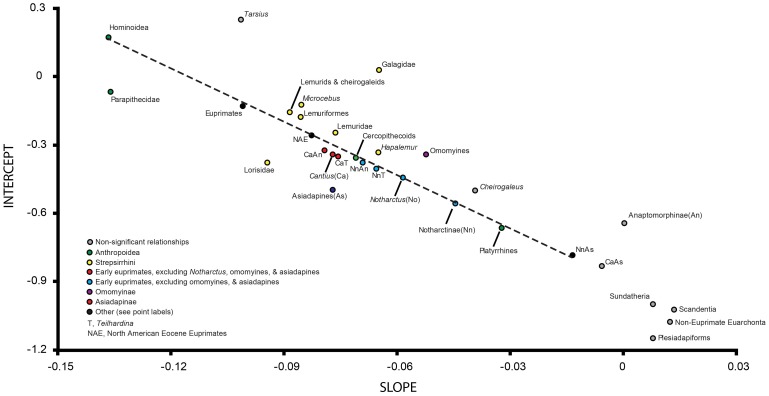
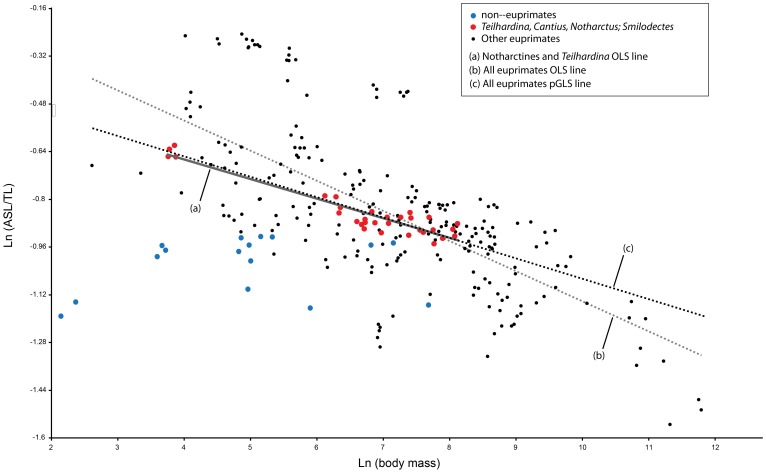
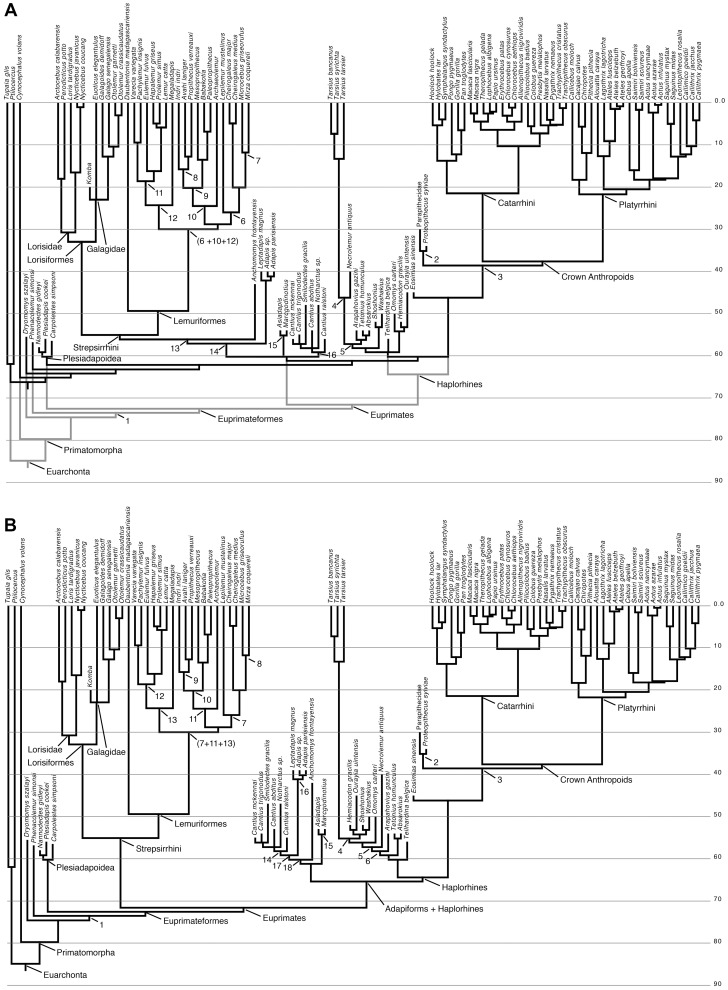
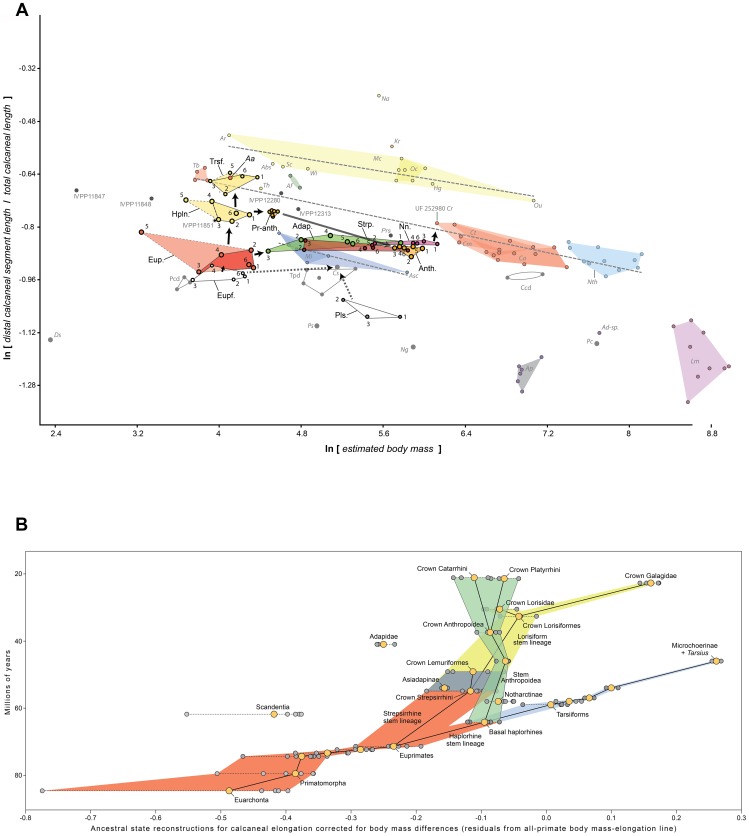
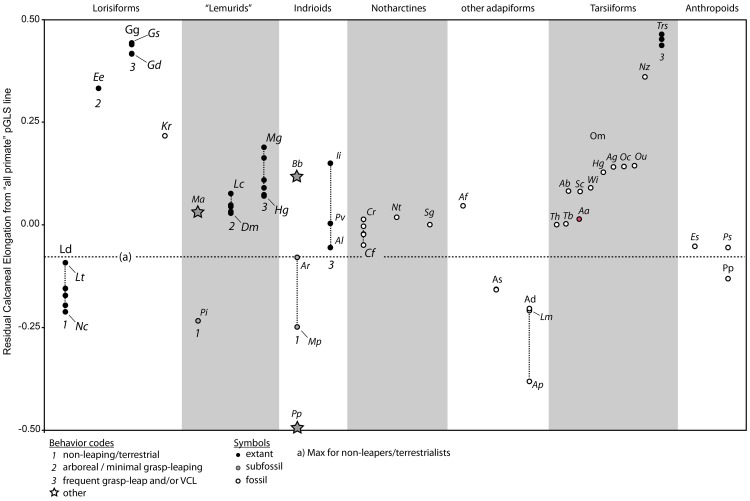
Specialized acrobatic leaping has been recognized as a key adaptive trait tied to the origin and subsequent radiation of euprimates based on its observed frequency in extant primates and inferred frequency in extinct early euprimates. Hypothesized skeletal correlates include elongated tarsal elements, which would be expected to aid leaping by allowing for increased rates and durations of propulsive acceleration at takeoff. Alternatively, authors of a recent study argued that pronounced distal calcaneal elongation of euprimates (compared to other mammalian taxa) was related primarily to specialized pedal grasping. Testing for correlations between calcaneal elongation and leaping versus grasping is complicated by body size differences and associated allometric affects. We re-assess allometric constraints on, and the functional significance of, calcaneal elongation using phylogenetic comparative methods, and present an evolutionary hypothesis for the evolution of calcaneal elongation in primates using a Bayesian approach to ancestral state reconstruction (ASR). Results show that among all primates, logged ratios of distal calcaneal length to total calcaneal length are inversely correlated with logged body mass proxies derived from the area of the calcaneal facet for the cuboid. Results from phylogenetic ANOVA on residuals from this allometric line suggest that deviations are explained by degree of leaping specialization in prosimians, but not anthropoids. Results from ASR suggest that non-allometric increases in calcaneal elongation began in the primate stem lineage and continued independently in haplorhines and strepsirrhines. Anthropoid and lorisid lineages show stasis and decreasing elongation, respectively. Initial increases in calcaneal elongation in primate evolution may be related to either development of hallucal-grasping or a combination of grasping and more specialized leaping behaviors. As has been previously suggested, subsequent increases in calcaneal elongation are likely adaptations for more effective acrobatic leaping, highlighting the importance of this behavior in early euprimate evolution.
Conflict of interest statement
Figures
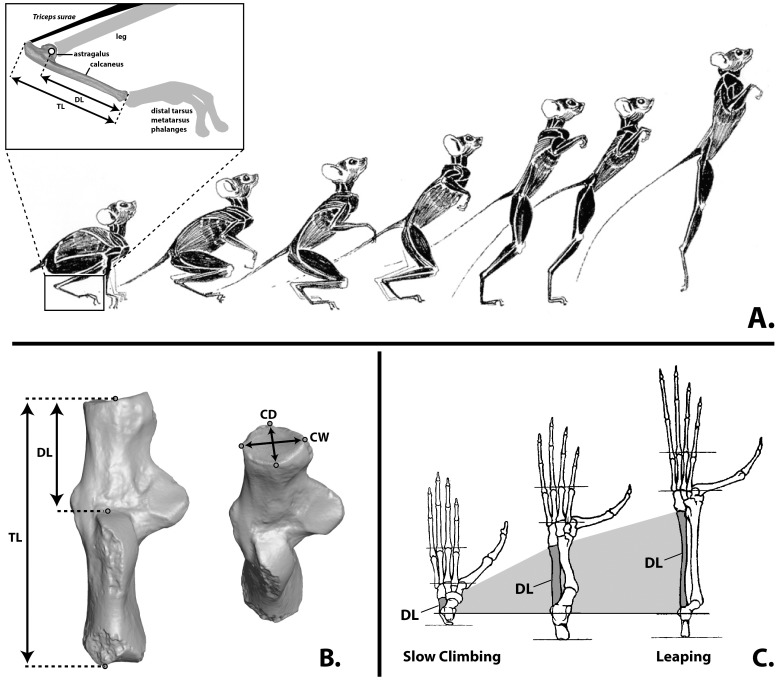
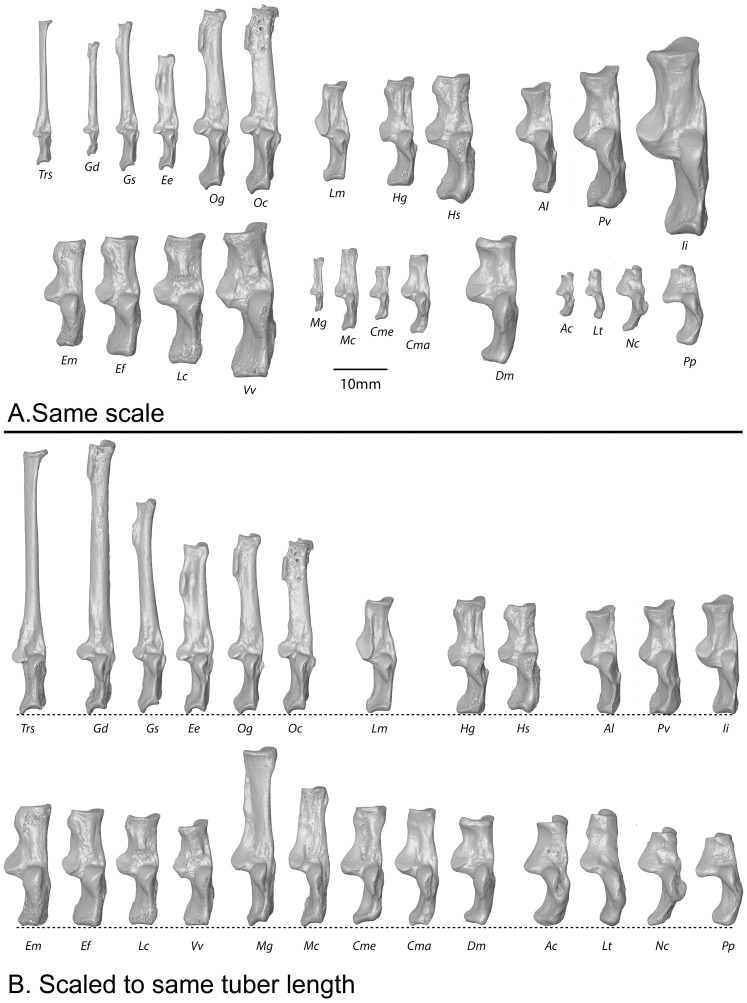
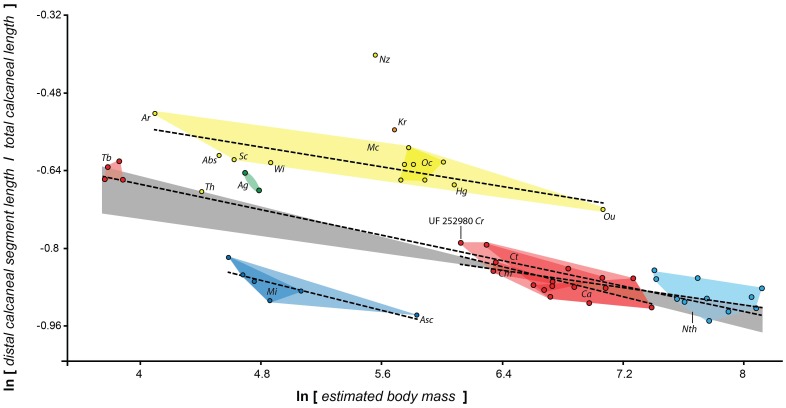
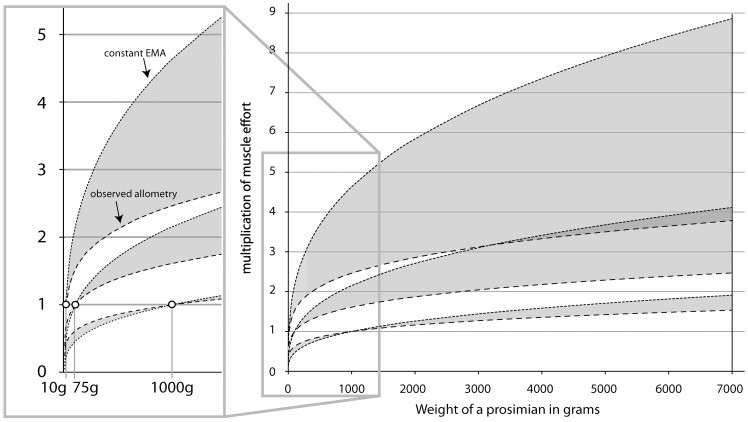
References
-
- Le Gros Clark WE (1959) The Antecedents of Man. Edinburgh: Edinburgh University Press. 394 p.
-
- Cartmill M (1992) New views on primate origins. Evol Anthropol 1: 105–111. - PubMed
-
- Szalay FS, Dagosto M (1980) Locomotor adaptations as reflected on the humerus of Paleogene primates. Folia Primatol 34: 1–45. - PubMed
-
- Crompton RH (1995) “Visual predation,” habitat structure, and the ancestral primate niche. In: Alterman L, Doyle GA, Izard MK, editors. Creatures of the Dark: The Nocturnal Prosimians. New York: Plenum Press. 11–30.
-
- Napier JR, Walker AC (1967) Vertical clinging and leaping - a newly recognized category of primate locomotion. Folia Primatol 6: 204–219. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
