

Age-related changes of myocardial ATP supply and demand mechanisms
- PMID: 23845538
- PMCID: PMC3783621
- DOI: 10.1016/j.tem.2013.06.001
Age-related changes of myocardial ATP supply and demand mechanisms
Abstract
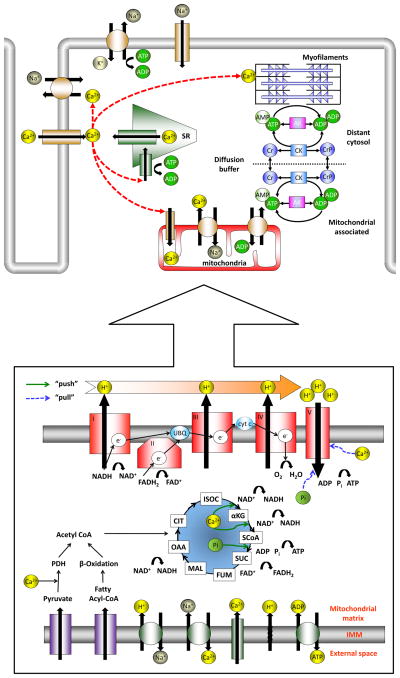
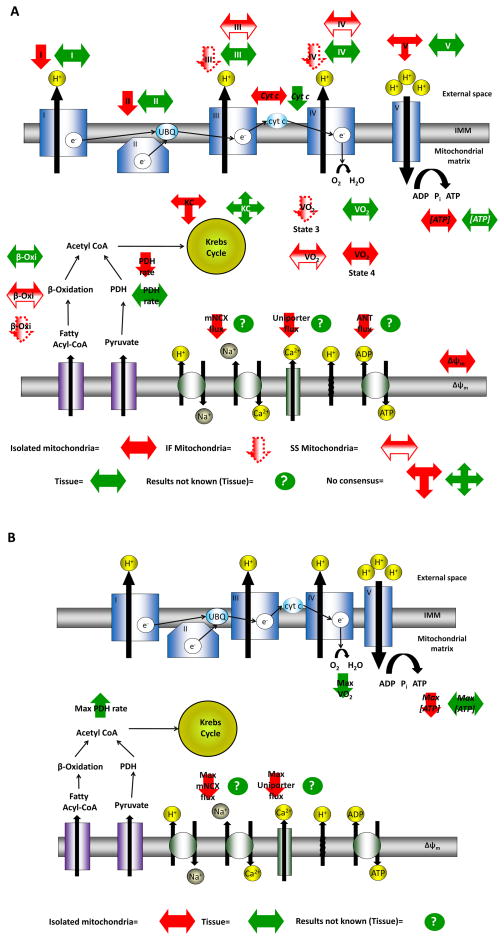
In advanced age, the resting myocardial oxygen consumption rate (MVO2) and cardiac work (CW) in the rat remain intact. However, MVO2, CW and cardiac efficiency achieved at high demand are decreased with age, compared to maximal values in the young. Whether this deterioration is due to decrease in myocardial ATP demand, ATP supply, or the control mechanisms that match them remains controversial. Here we discuss evolving perspectives of age-related changes of myocardial ATP supply and demand mechanisms, and critique experimental models used to investigate aging. Specifically, we evaluate experimental data collected at the level of isolated mitochondria, tissue, or organism, and discuss how mitochondrial energetic mechanisms change in advanced age, both at basal and high energy-demand levels.
Keywords: aging; bioenergetics; cardiac work; mitochondria; respiration.
Published by Elsevier Ltd.
Figures
References
-
- van Beek JH. Adenine nucleotide-creatine-phosphate module in myocardial metabolic system explains fast phase of dynamic regulation of oxidative phosphorylation. Am J Physiol Cell Physiol. 2007;293:C815–829. - PubMed
-
- Saks VA, et al. Functional coupling as a basic mechanism of feedback regulation of cardiac energy metabolism. Mol Cell Biochem. 2004;256–257:185–199. - PubMed
-
- Bak MI, et al. Interaction of hypoxia and aging in the heart: analysis of high energy phosphate content. J Mol Cell Cardiol. 1998;30:661–672. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
