

The transverse-axial tubular system of cardiomyocytes
- PMID: 23846763
- PMCID: PMC11113601
- DOI: 10.1007/s00018-013-1410-5
The transverse-axial tubular system of cardiomyocytes
Abstract
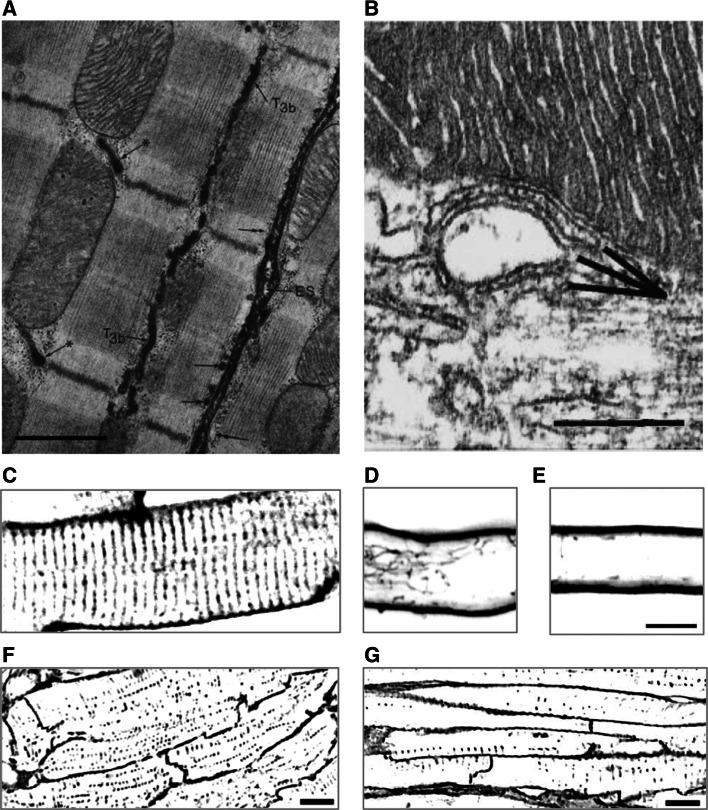
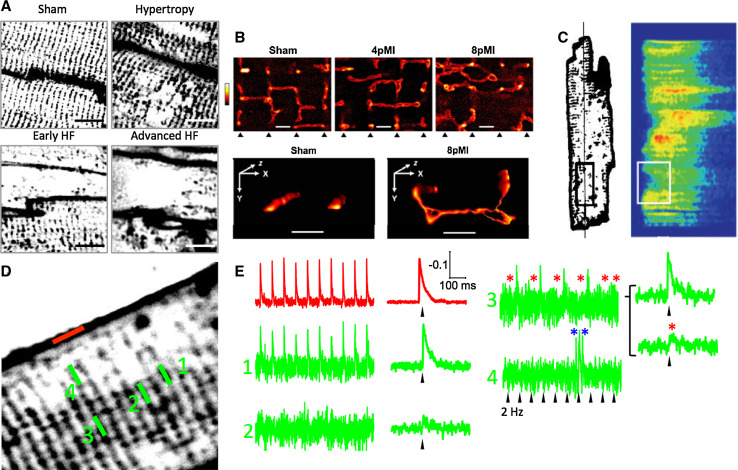
A characteristic histological feature of striated muscle cells is the presence of deep invaginations of the plasma membrane (sarcolemma), most commonly referred to as T-tubules or the transverse-axial tubular system (TATS). TATS mediates the rapid spread of the electrical signal (action potential) to the cell core triggering Ca(2+) release from the sarcoplasmic reticulum, ultimately inducing myofilament contraction (excitation-contraction coupling). T-tubules, first described in vertebrate skeletal muscle cells, have also been recognized for a long time in mammalian cardiac ventricular myocytes, with a structure and a function that in recent years have been shown to be far more complex and pivotal for cardiac function than initially thought. Renewed interest in T-tubule function stems from the loss and disorganization of T-tubules found in a number of pathological conditions including human heart failure (HF) and dilated and hypertrophic cardiomyopathies, as well as in animal models of HF, chronic ischemia and atrial fibrillation. Disease-related remodeling of the TATS leads to asynchronous and inhomogeneous Ca(2+)-release, due to the presence of orphan ryanodine receptors that have lost their coupling with the dihydropyridine receptors and are either not activated or activated with a delay. Here, we review the physiology of the TATS, focusing first on the relationship between function and structure, and then describing T-tubular remodeling and its reversal in disease settings and following effective therapeutic approaches.
Figures




References
-
- Lindner E. Submicroscopic morphology of the cardiac muscle. Z Zellforsch Mikrosk Anat. 1957;45(6):702–746. - PubMed
-
- Tidball JG, Cederdahl JE, Bers DM. Quantitative analysis of regional variability in the distribution of transverse tubules in rabbit myocardium. Cell Tissue Res. 1991;264(2):293–298. - PubMed
-
- Franzini-Armstrong C, Venosa RA, Horowicz P. Morphology and accessibility of the ‘transverse’ tubular system in frog sartorius muscle after glycerol treatment. J Membr Biol. 1973;14(3):197–212. - PubMed
-
- Bers DM. Cardiac excitation-contraction coupling. Nature. 2002;415(6868):198–205. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
