

Mitochondrial respiration without ubiquinone biosynthesis
- PMID: 23847050
- PMCID: PMC3888124
- DOI: 10.1093/hmg/ddt330
Mitochondrial respiration without ubiquinone biosynthesis
Abstract
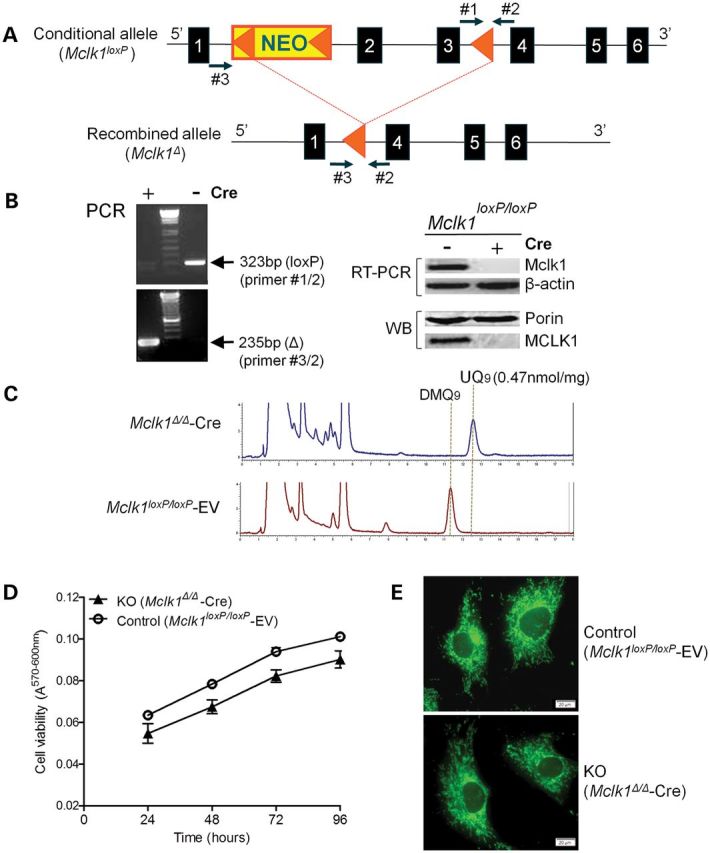
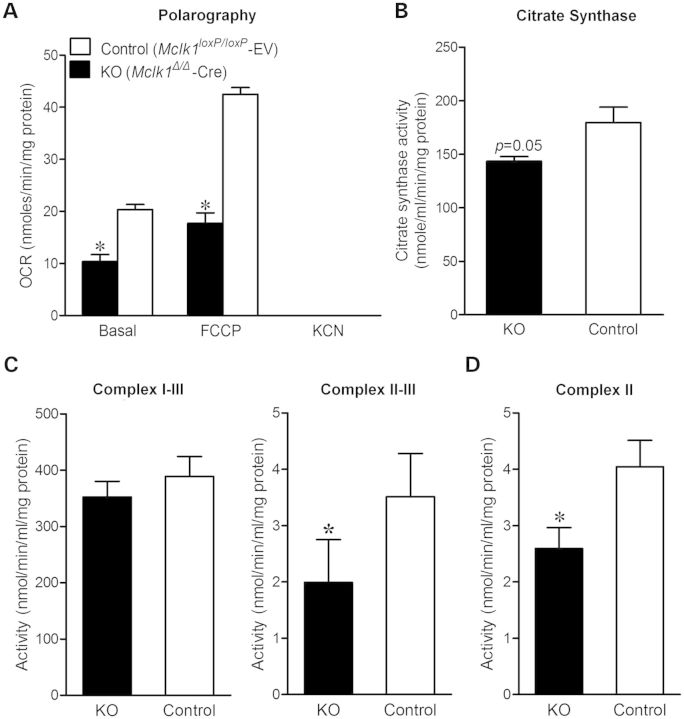
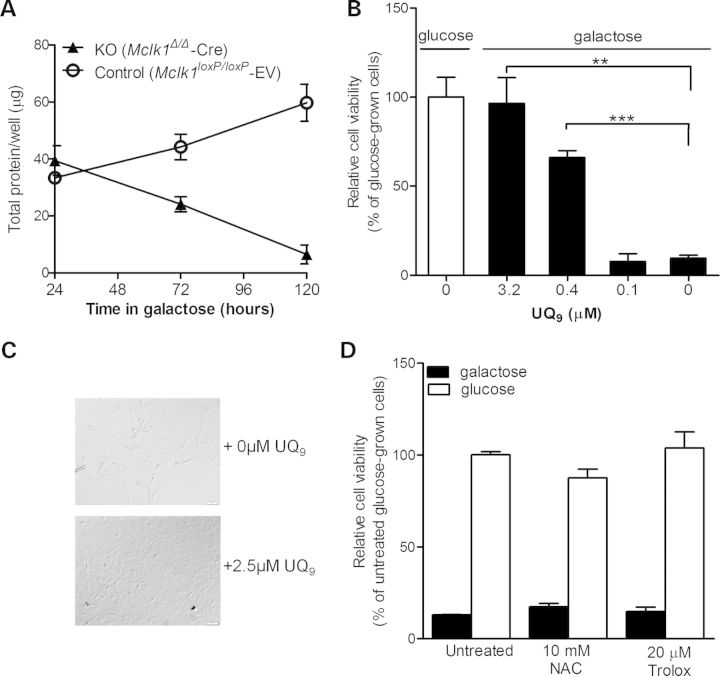
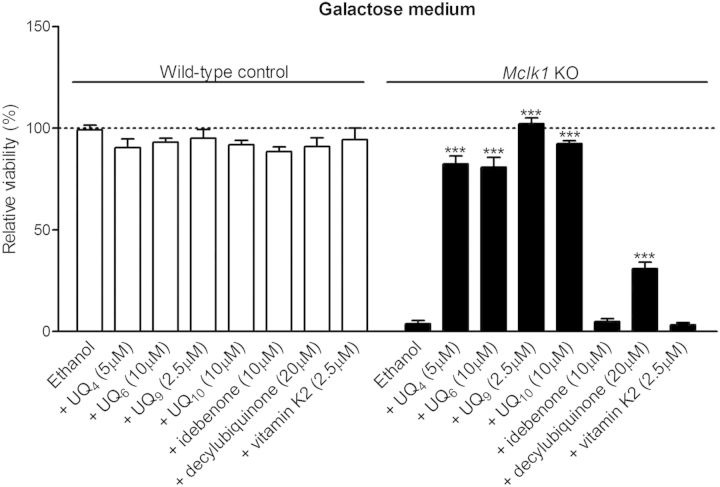
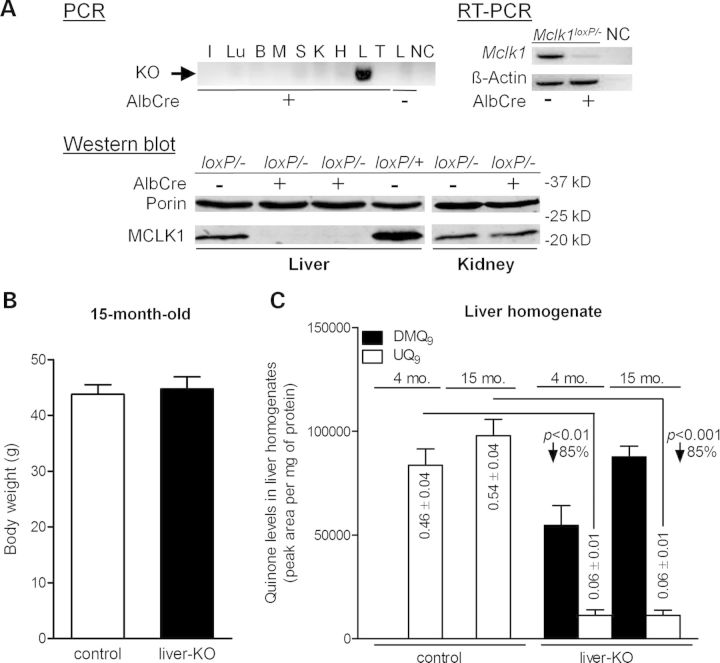
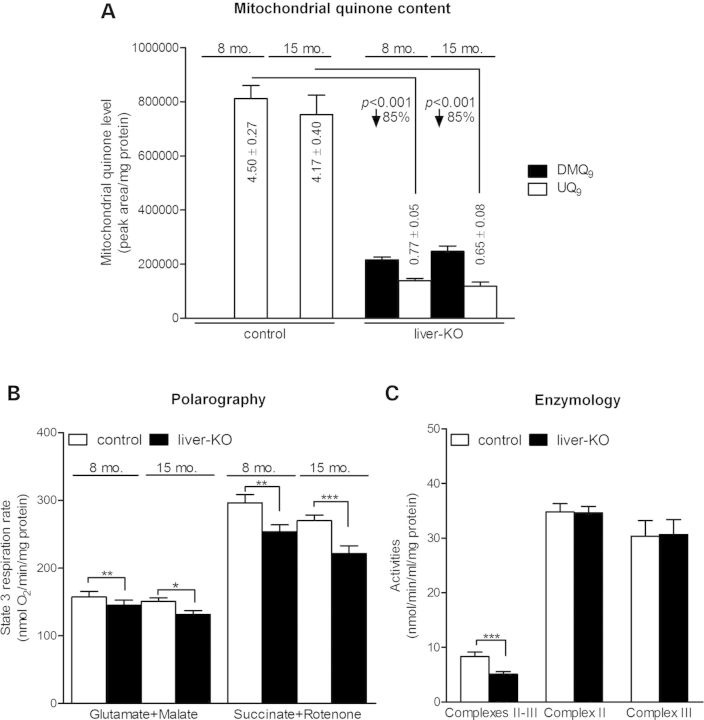
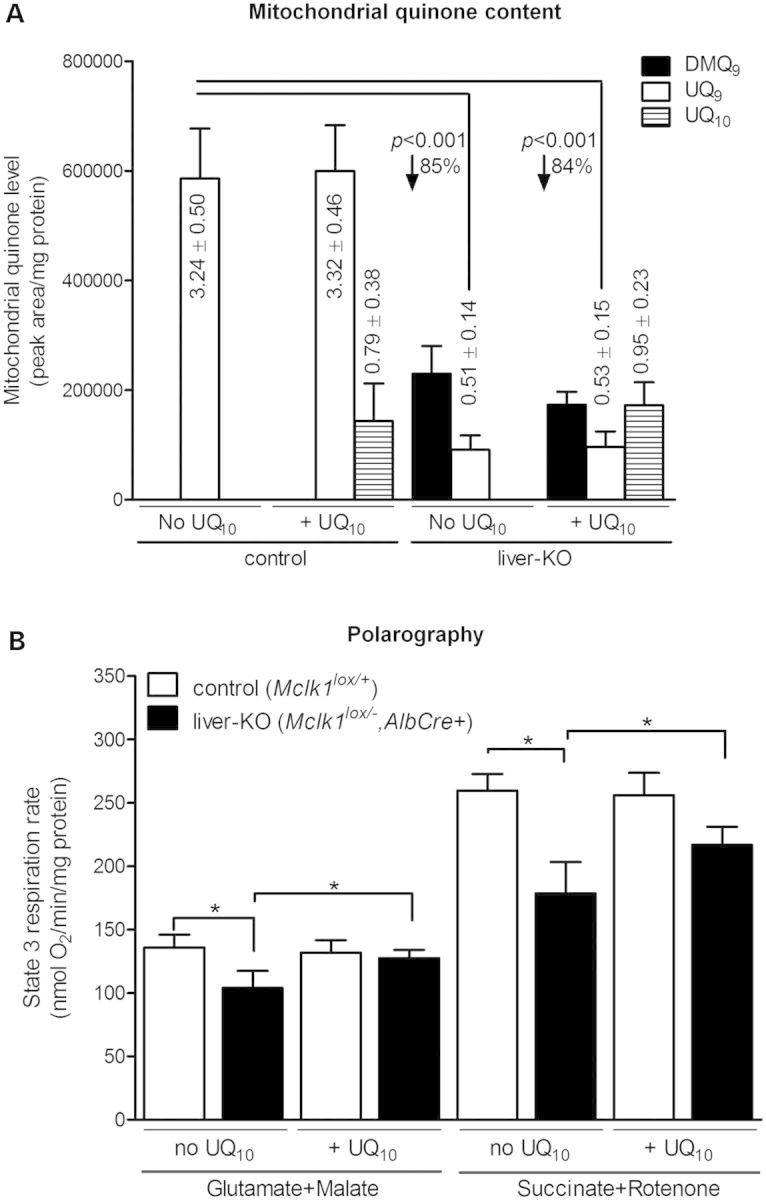
Ubiquinone (UQ), a.k.a. coenzyme Q, is a redox-active lipid that participates in several cellular processes, in particular mitochondrial electron transport. Primary UQ deficiency is a rare but severely debilitating condition. Mclk1 (a.k.a. Coq7) encodes a conserved mitochondrial enzyme that is necessary for UQ biosynthesis. We engineered conditional Mclk1 knockout models to study pathogenic effects of UQ deficiency and to assess potential therapeutic agents for the treatment of UQ deficiencies. We found that Mclk1 knockout cells are viable in the total absence of UQ. The UQ biosynthetic precursor DMQ9 accumulates in these cells and can sustain mitochondrial respiration, albeit inefficiently. We demonstrated that efficient rescue of the respiratory deficiency in UQ-deficient cells by UQ analogues is side chain length dependent, and that classical UQ analogues with alkyl side chains such as idebenone and decylUQ are inefficient in comparison with analogues with isoprenoid side chains. Furthermore, Vitamin K2, which has an isoprenoid side chain, and has been proposed to be a mitochondrial electron carrier, had no efficacy on UQ-deficient mouse cells. In our model with liver-specific loss of Mclk1, a large depletion of UQ in hepatocytes caused only a mild impairment of respiratory chain function and no gross abnormalities. In conjunction with previous findings, this surprisingly small effect of UQ depletion indicates a nonlinear dependence of mitochondrial respiratory capacity on UQ content. With this model, we also showed that diet-derived UQ10 is able to functionally rescue the electron transport deficit due to severe endogenous UQ deficiency in the liver, an organ capable of absorbing exogenous UQ.
Figures
Similar articles
-
The submitochondrial distribution of ubiquinone affects respiration in long-lived Mclk1+/- mice.J Cell Biol. 2012 Oct 15;199(2):215-24. doi: 10.1083/jcb.201203090. Epub 2012 Oct 8. J Cell Biol. 2012. PMID: 23045551 Free PMC article.
-
The idebenone metabolite QS10 restores electron transfer in complex I and coenzyme Q defects.Biochim Biophys Acta Bioenerg. 2018 Sep;1859(9):901-908. doi: 10.1016/j.bbabio.2018.04.006. Epub 2018 Apr 22. Biochim Biophys Acta Bioenerg. 2018. PMID: 29694828
-
The molecular genetics of coenzyme Q biosynthesis in health and disease.Biochimie. 2014 May;100:78-87. doi: 10.1016/j.biochi.2013.12.006. Epub 2013 Dec 16. Biochimie. 2014. PMID: 24355204 Review.
-
Ubiquinone is necessary for mouse embryonic development but is not essential for mitochondrial respiration.J Biol Chem. 2001 Dec 7;276(49):46160-4. doi: 10.1074/jbc.M108980200. Epub 2001 Oct 3. J Biol Chem. 2001. PMID: 11585841
-
Molecular genetics of ubiquinone biosynthesis in animals.Crit Rev Biochem Mol Biol. 2013 Jan-Feb;48(1):69-88. doi: 10.3109/10409238.2012.741564. Epub 2012 Nov 29. Crit Rev Biochem Mol Biol. 2013. PMID: 23190198 Free PMC article. Review.
Cited by
-
Resveratrol and para-coumarate serve as ring precursors for coenzyme Q biosynthesis.J Lipid Res. 2015 Apr;56(4):909-19. doi: 10.1194/jlr.M057919. Epub 2015 Feb 14. J Lipid Res. 2015. PMID: 25681964 Free PMC article.
-
CoQ10 supplementation rescues nephrotic syndrome through normalization of H2S oxidation pathway.Biochim Biophys Acta Mol Basis Dis. 2018 Nov;1864(11):3708-3722. doi: 10.1016/j.bbadis.2018.09.002. Epub 2018 Sep 6. Biochim Biophys Acta Mol Basis Dis. 2018. PMID: 30251690 Free PMC article.
-
How plants synthesize coenzyme Q.Plant Commun. 2022 Sep 12;3(5):100341. doi: 10.1016/j.xplc.2022.100341. Epub 2022 May 25. Plant Commun. 2022. PMID: 35614856 Free PMC article. Review.
-
Animal Models of Coenzyme Q Deficiency: Mechanistic and Translational Learnings.Antioxidants (Basel). 2021 Oct 26;10(11):1687. doi: 10.3390/antiox10111687. Antioxidants (Basel). 2021. PMID: 34829558 Free PMC article. Review.
-
Border between natural product and drug: comparison of the related benzoquinones idebenone and coenzyme Q10.Redox Biol. 2015;4:289-95. doi: 10.1016/j.redox.2015.01.009. Epub 2015 Jan 14. Redox Biol. 2015. PMID: 25625583 Free PMC article. Review.
References
-
- Crane F.L. Discovery of ubiquinone (coenzyme Q) and an overview of function. Mitochondrion. 2007;7(suppl):S2–S7. - PubMed
-
- Lenaz G. A critical appraisal of the mitochondrial coenzyme Q pool. FEBS Lett. 2001;509:151–155. - PubMed
-
- Bentinger M., Brismar K., Dallner G. The antioxidant role of coenzyme Q. Mitochondrion. 2007;7(suppl):S41–S50. - PubMed
-
- Walter L., Miyoshi H., Leverve X., Bernard P., Fontaine E. Regulation of the mitochondrial permeability transition pore by ubiquinone analogs. A progress report. Free Rad. Res. 2002;36:405–412. - PubMed
-
- Evans D.R., Guy H.I. Mammalian pyrimidine biosynthesis: fresh insights into an ancient pathway. J. Biol. Chem. 2004;279:33035–33038. - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
