

Systems approach to developmental biology--designs for robust patterning
- PMID: 23847812
- PMCID: PMC8687163
- DOI: 10.1049/iet-syb.2012.0042
Systems approach to developmental biology--designs for robust patterning
Abstract
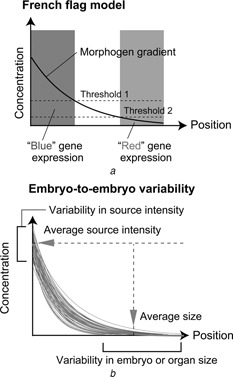
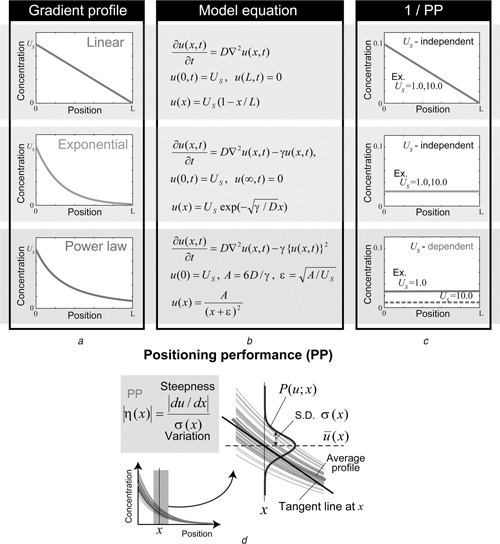
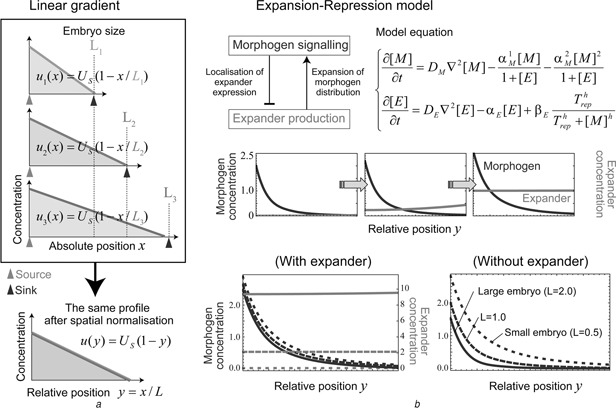
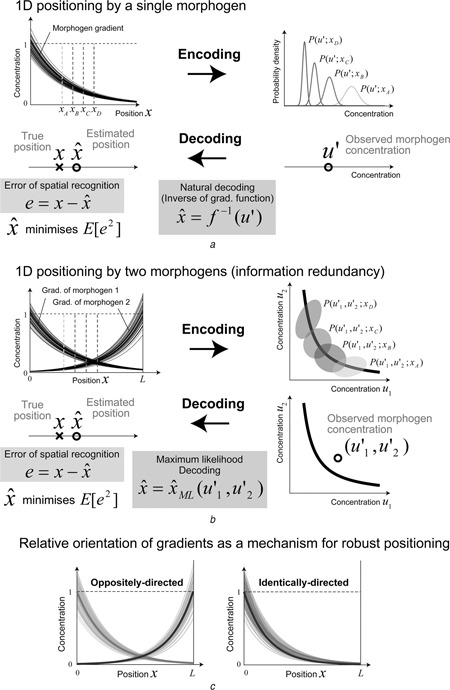
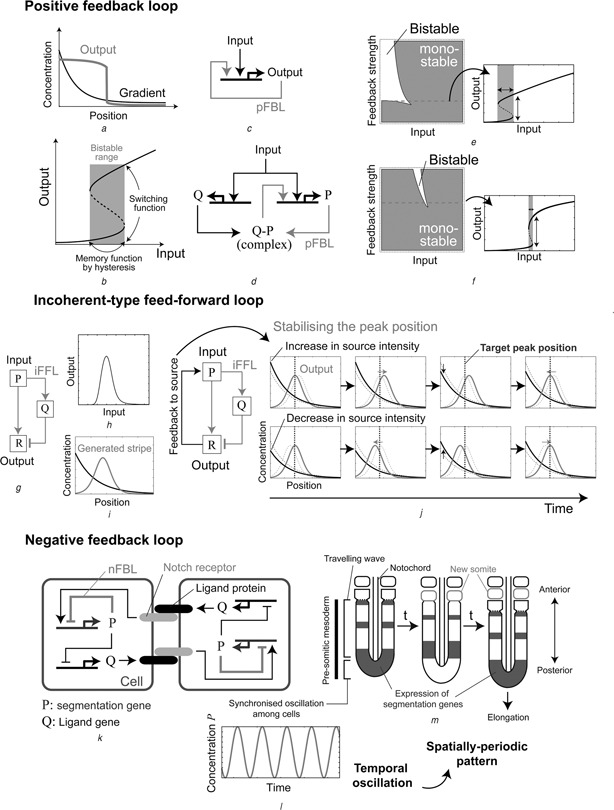
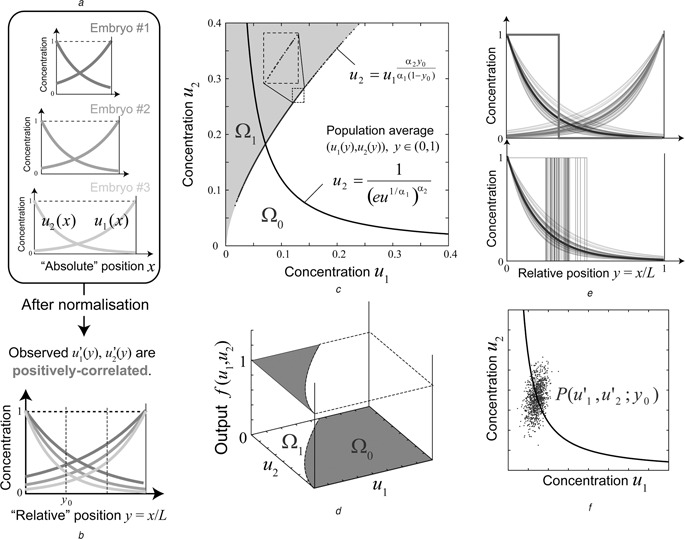
Patterning is an important step in animal development that generates spatially non-uniform gene expression patterns or spatially heterogeneous cellular responses. Patterning is realised by the generation and reading of positional information provided by spatial gradients of morphogens, diffusive chemicals in the extracellular environment. To achieve normal development, accurate patterning that is robust against noise is necessary. Here the authors describe how morphogen gradient formation and gradient interpretation processes are designed to achieve highly reproducible patterning. Furthermore, recent advancements in measurement and imaging techniques have enabled researchers to obtain quantitative dynamic and multi-physical data, not only for chemical events, but also for the geometrical and mechanical properties of cells in vivo. The authors briefly review some recent studies on the effects of such non-chemical events on patterning.
Figures

Similar articles
-
Morpheus unbound: reimagining the morphogen gradient.Cell. 2007 Jan 26;128(2):245-56. doi: 10.1016/j.cell.2007.01.004. Cell. 2007. PMID: 17254964 Review.
-
Spatiotemporal mechanisms of morphogen gradient interpretation.Curr Opin Genet Dev. 2011 Dec;21(6):726-31. doi: 10.1016/j.gde.2011.10.002. Epub 2011 Oct 25. Curr Opin Genet Dev. 2011. PMID: 22033220 Free PMC article. Review.
-
Mathematical model of the formation of morphogen gradients through membrane-associated non-receptors.Bull Math Biol. 2010 May;72(4):805-29. doi: 10.1007/s11538-009-9470-2. Epub 2009 Oct 31. Bull Math Biol. 2010. PMID: 19882189
-
Steady-state invariant genetics: probing the role of morphogen gradient dynamics in developmental patterning.J R Soc Interface. 2011 Oct 7;8(63):1429-39. doi: 10.1098/rsif.2010.0738. Epub 2011 Mar 18. J R Soc Interface. 2011. PMID: 21421746 Free PMC article.
-
Encoding and decoding of positional information in morphogen-dependent patterning.Curr Opin Genet Dev. 2012 Dec;22(6):553-61. doi: 10.1016/j.gde.2012.10.002. Epub 2012 Nov 28. Curr Opin Genet Dev. 2012. PMID: 23200115 Review.
Cited by
-
Top-down models in biology: explanation and control of complex living systems above the molecular level.J R Soc Interface. 2016 Nov;13(124):20160555. doi: 10.1098/rsif.2016.0555. J R Soc Interface. 2016. PMID: 27807271 Free PMC article. Review.
-
Re-membering the body: applications of computational neuroscience to the top-down control of regeneration of limbs and other complex organs.Integr Biol (Camb). 2015 Dec;7(12):1487-517. doi: 10.1039/c5ib00221d. Epub 2015 Nov 16. Integr Biol (Camb). 2015. PMID: 26571046 Free PMC article. Review.
References
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
